

MAPKカスケード
MAPK(mitogen-activated protein kinase)はリン酸化酵素(セリン/スレオニンキナーゼ)の一つで、特定のタンパク質をリン酸化して活性化させますが、自分の活性化に自身のリン酸化が必要なため、MAPKK(MAPK kinase)がMAPKをリン酸化することが必要です。
さらに、MAPKKはMAPKKKによって、MAPKKKはMAPKKKKによってリン酸化されることで活性化されます(Kがたくさん続くのでKKKは3Kのように略されることあり)。
MAPKは何も刺激がない状態では細胞質に存在し、刺激があるとMAPKKとの結合がはずれて核内へと進入しますが、これには核外移行シグナル(NES)が大きくかかわっています。 MAPKKがNESを持っているために、結合している状態では核内に移行できません。
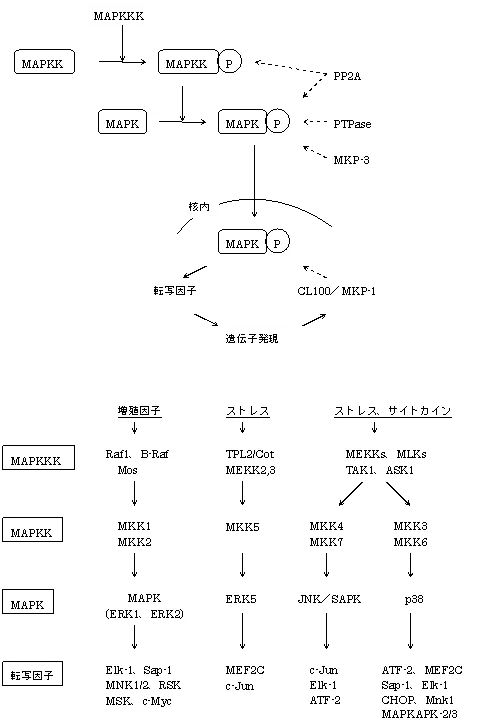
このようなMAPKカスケードはMAPKの種類によって経路が大きく4つあります。
一つは、Rasの下流で起こる古典的MAPK経路と呼ばれるもので、Raf-1がMAP3Kとして働きMAP2KであるMEK1/2をリン酸化して、ERK1/2が活性化するルートです。 この経路は増殖因子の刺激で通るルートでAP-1の構成因子:c-fosやアラキドン酸カスケードを進行させるcPLA2(ホスホリパーゼA2)を作るのに必須の経路です。
二つめは、TNFαやIL-1がTRAF2を介してMEKK1(MAP3K)→MKK4/7(MAP2K)→JNK1/2/3/SAPK(MAPK)と進んで、AP-1の構成因子:c-junを作るルートです。
三つめは、ストレスから活性化される経路で、TNFRやIL-1RからTRAF2→ASK1(MAP3K)、またはFasやTGF-βからDAXX→ASK1と進んでp38α/β/γ/δが活性化されるルートです。
四つめは、あまりよく知られていませんが、ERK5経路と呼ばれ、これも酸化ストレス、EGFなどの一部の増殖因子で活性化し、増殖に関与するといわれる。
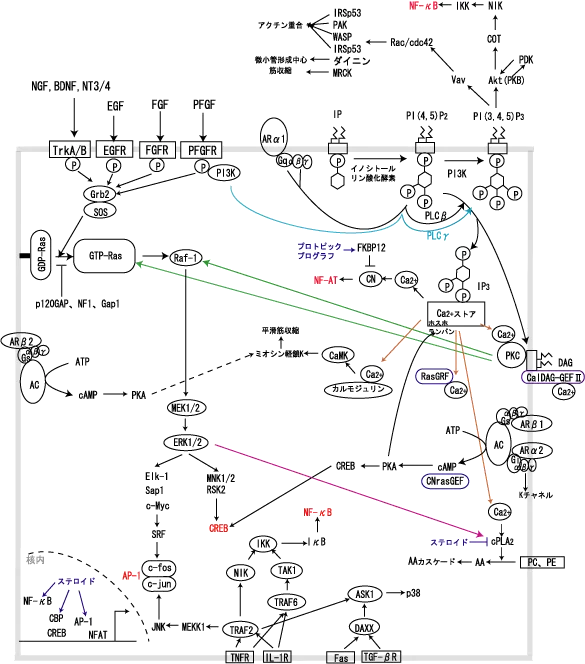
さて、次にMAPKカスケードの周りのシグナルを詳しく見ていきましょう。
まず一番こんがらがってくるところは、イノシトールリン脂質あたりでしょうか?勝手にそう決め付けて話を進めます。。。
イノシトールリン脂質というのは、細胞膜のリン脂質に組み込まれた、グリセロール+脂肪酸+リン酸+イノシトールからなるタンパク質であり、リン酸が一つだけ付いている IP(ホスファチジルイノシトール)はRasの下流で活性化されるイノシトールリン酸化酵素によって4位と5位のリン酸化が起きて、PI(4,5)P2へと変わる。
PI(4,5)P2は、PLC(ホスホリパーゼ)によってDAG(ジアシルグリセロール)とIP3(イノシトール3リン酸)に分解され、IP3は筋小胞体のCaストアからCaの放出を促進させ、 DAGはそのCaと共にPKC(プロテインキナーゼC)に結合することで、PKCが活性化されRasやRafのリン酸化による活性化に作用する。
PLCにもいくつかのサブタイプがありますが、重要なのはPLCβとPLCγの二つで、PLCβはアドレナリンα1受容体シグナルがGqα/β/γタンパク質を活性化することで活性化され、 PLCγはPI3Kなどの下流で活性化されます(PLCγはSH2、SH3ドメインを持つ)。
また、プロテインキナーゼにもA、B、Cと3つのサブタイプがあります。
PKAは、βやα2受容体からのAC(アデニル酸シクラーゼ)→cAMP(サイクリックAMP)産生にて活性化され、cAMPにより活性化されるCREB(cAMP response element-binding protein)は、CBPなどの転写調節因子の補佐の元、CREに結合して転写を促進する。 PKBは、Aktの別名で、MAP3KであるCOTをリン酸化して、NIK→NF-κB活性化という経路に必要です。 PKCは、RasやRafをリン酸化することで細胞増殖を活性化します。PKC自体にもα,β,γなどいくつかのサブタイプがありますがここでは述べません。
話は戻り、PI(4,5)P2からの経路ですが、このようにPLCによって分解されるルートのほかに、PI3K(phosphatidylinositol-3-kinase)という酵素によってさらにリン酸化される経路があります。
PI3KはPI(4,5)P2の3位にさらにリン酸を付加して、PI(3,4,5)P3を作ります。 PI3KはSH2ドメインを持った調節サブユニットと触媒サブユニットを持ち、このリン酸化作用以外にも様々な基質のチロシンリン酸化部位、アダプター分子やドッキングタンパク質 などと結合してシグナルを伝達するアダプター分子的な作用を持っています。
PI(3,4,5)P3は、PHドメインを持つタンパク質をリクルートして活性化する作用を持っています。このPHドメインを有する活性化されるタンパク質の代表として、VavとAktが上げられます。
Akt(PKB)はキナーゼとPHドメインを持つタンパク質で、AktのPHドメインでPI(3,4,5)P3に結合すると、蛋白リン酸化酵素PDK(phosphoinositide dependent protein kinase)の基質になり 、PDKからリン酸化を受けれるようになる。 PDKにリン酸化されたAktは、COT→NIK→とすすんでアポトーシスを抑制する。
VavはGEFの仲間でこれもPHドメインを持っている。PI(3,4,5)P3にリクルートされたVavがリン酸化されると、GEFとして、GDP-Racを活性型のGTP-Racへと変換する。
RacやCdc42はSmall Gタンパク質と呼ばれるタンパク質の中でもRhoファミリーに属しています。 Small Gタンパク質にはRas、Rho、Rab、Arfなどがあり、三量体Gタンパク質とは違って、単量体で働くところは違いますが、GDP結合型が不活性で、GTP結合型が活性であるところは共通です。
Small Gタンパク質のGDPをはずしてGTPをくっつける因子をGEF(guanine nucleotide exchange factor)と呼び、GTPをはずしてGDPをくっつける因子をGAP(GTPase activating protein) と呼びます。
上記の図の中で青い枠で囲ったもの3種(CalDAG-GEFⅡ、CNrasGEF、RasGRF)はGEFに属します。活性化したRasを不活化するGAPとしてはp120GAP、NF1、Gap1が存在する。NF1は静止期のRasを不活性の状態に保つ、p120GAPとNF1は活性かされたRasを不活性な状態に戻す。Gap1はGタンパク質により活性化され、Rasシグナル伝達を抑制する。
一方、Rac、Cdc42などのRhoが活性化されると、アクチン細胞骨格系(p21G蛋白であるRhoとその基質であるROCK(Rho-associated coiled-coil forming protein kinase)による系) が制御される。なお、SOSおよびRasGRFはRhoファミリーのRacを直接活性化する。
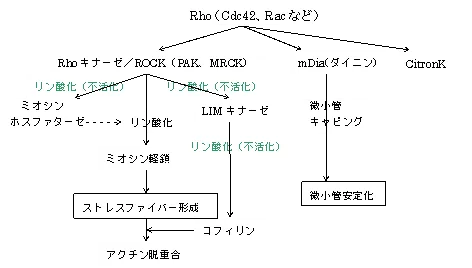
ストレスファイバーというのは、別名緊張繊維と呼ばれ、アクチンフィラメントが数十本束化したものを言います。アトピー性皮膚炎では、EGF、NGFなど の成長因子が多量に出される上に、交感神経緊張の状態、つまりGタンパク質の活性化が起こるので、Rasが健常人に比べて活性化されています。
そのため、細胞間脂質の量は少なくても、細胞の骨格自体は安定化します。
- DNAと遺伝子とは
- DNAの転写
- タンパク質の合成
- 初期mRNAを作るまで
- チロシンキナーゼ型受容体
- JAK-STAT型受容体
- TGF受容体
- TNF受容体
- Wnt受容体
- Notch受容体
- Toll-like受容体
- NOD-like受容体
- RIG-like受容体
- MAPKカスケード
- タンパク質の分解
- 癌抑制遺伝子p53
- CKIとは?
- 細胞周期
- DNAの複製
コメントor補足情報orご指摘あればをお願いします。
- << 前のページ
- 次のページ >>
