

ドパミン神経系
ドパミンの合成
ドパミンは、チロシン(必須アミノ酸ではない)からチロシン水酸化酵素によってL-ドパへ、L-ドパはドパ脱炭酸酵素によってドパミンに合成される。
ドパミンはドパミンβ水酸化酵素の働きで、ノルアドレナリンにも合成される。ノルアドレナリンはCOMT(カテコール-O-メチル転移酵素)によってノルメタネフリンへ、MAO(モノアミン酸化酵素)によって、MHPG(3-メトキシ-4水酸化フェニールグリコール)へと分解され代謝される。
ドパミンの神経核
ドパミンの神経群として重要なのが、A9:黒質緻密部とA10:腹側被蓋野の2つである。
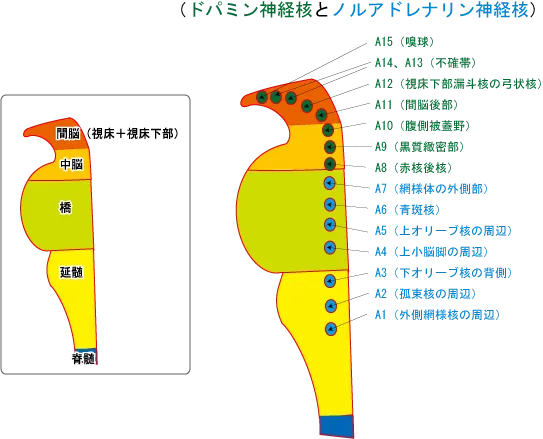
| 神経集団番号 | 神経集団名称 | 主な作用 |
|---|---|---|
| A8 | 赤核後核 | 辺縁系へ投射 |
| A9 | 黒質緻密部 | 新線条体へ投射。パーキンソン病の関連部位として錐体外路運動に関与 |
| A10 | 腹側被蓋野 | 辺縁系へ投射する中脳辺縁系路と前頭葉へ投射する中脳皮質路がある。前者は扁桃体の興奮(情動)により活性化され、後者はストレスや不安等負の要因で活性化。 |
| A11 | 間脳後部 | 視床下部へ投射。レストレスレッグス症候群の関連部位 |
| A12 | 弓状核 | 下垂体ホルモンの分泌に制御する。この系の抑制はプロラクチンの分泌を促進する(D2ブロッカーの高プロラクチン血症の原因部位)。 |
| A13、A14 | 不確帯 | 視床下部へ投射 |
| A15 | 嗅球 |
ドパミンの作用
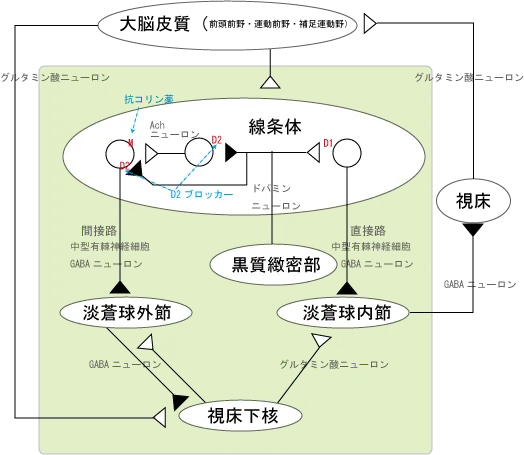
A9:黒質-線条体路
黒質-線条体路では、中脳の黒質に細胞体を置くドパミンニューロンが、線条体でGABAを伝達物質とする抑制性の中型有棘神経細胞を抑制的に制御しています。
この抑制制御がうまく行かない上に、アセチルコリン神経の活性化によりGABA神経の興奮→視床下核の興奮→視床の抑制→錐体外路障害を生じるのがパーキンソン病となります。
統合失調症では、主として前頭葉のNMDA受容体やドパミン受容体の異常によってドパミンの分泌が亢進し、ここ(基底核)のD2受容体を刺激してしまうため、視床が興奮して、ちょっとのことでもビクッとしてしまうような知覚過敏の状態を作り出してしまいます。
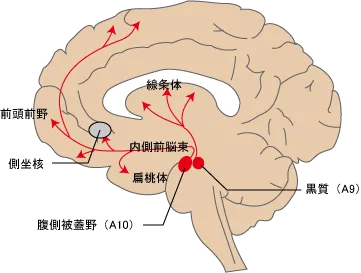
A10:腹側被蓋野(中脳辺縁系路・中脳皮質路)
中脳腹側被蓋野から投射されるドパミンニューロンは大きく、腹側被蓋野-辺縁系路(側坐核・海馬・扁桃体等)と腹側被蓋野-皮質路(前頭前野)の2つに分けることができます。
中脳辺縁系路は正の強化因子、快情動、報酬系に関わる情動回路を動かすドパミンを分泌する経路、統合失調症では陽性症状の発現に関わっています。
さすってもらったら痛みが収まった→快情報として扁桃体が認識→記憶として海馬へ格納。この快の記憶が原因で扁桃体がA10を活性化→A10活性化によるドパミンが側坐核shellを興奮させて、視床内背側核→前頭前野→というYakovlevの情動回路を作動させて、もういちどさすって貰いたい!さすってもらえば痛みが収まる!といった快の情動行動を起こす方向へと進む。
中脳皮質路のほうは、不安やストレスにより活性化し、負の行動を起こす陰性症状の発現に関わっています。
ストレス刺激がA10経路を活性化すると、前頭前野のドパミンD1受容体を介して前頭前野の行う認知・思考・ワーキングメモリ等の機能を抑制してしまいます。
ドパミンの自己受容体
D2受容体には、D2L受容体とD2S受容体のサブタイプが存在している。
D2L受容体は、辺縁系では後シナプス、線条体では中型有棘細胞の後シナプスに多く発現している。D2S受容体は自己受容体として黒質、腹側被蓋野のドパミン神経終末(前シナプス)に発現している。
一方で、中脳皮質路のドパミン神経においては自己受容体が欠如しているため、ストレス刺激によって活性化してしまうとドパミンの分泌が制御できなくなるという。
- 参考・引用元
-
神経路の理解
├ 神経の分類
├ 脳、辺縁系、基底核
├ 視床と視床下部
├ 中脳
├ 橋
├ 延髄
├ 運動路
├ 体性感覚路(体・四肢)
├ 体性感覚路(顔)
├ 視覚路
├ 聴覚路
├ 前庭感覚路
├ 上行性網様体賦活系
├ 下行性抑制系
└ 反射系
体性神経(運動・感覚神経)
├ かゆみの種類・・・中枢性のかゆみと末梢性のかゆみ
├ かゆみの伝達の概略
├ 神経線維の種類・・・かゆみを伝える神経とは
├ かゆみ受容器・・・かゆみ刺激を受け取る受容器
├ かゆみ受容体・・・TRPファミリーを中心に
├ 痒みと痛みの関係
├ 興奮伝導・・・かゆみの伝導メカニズム
├ DRGとガッセル神経節・・・神経節に存在するもの(NGFなど)
└ 脊髄後角と尾側亜核・・・一次ニューロンから二次ニューロンへ
自律神経(交感・副交感神経)
├ 自律神経というもの
├ 交感神経
└ 副交感神経
中枢神経(NA・Ad・DA・5HT・Ach・GABA)
├ ノルアドレナリン
├ アドレナリン
├ ドパミン
├ セロトニン
├ アセチルコリン
└ γアミノ酪酸
コメントor補足情報orご指摘あればをお願いします。
- << 前のページ
- 次のページ >>
