

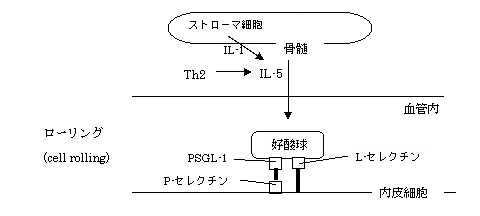
好酸球の遊走
好酸球はおもにIL-5の刺激を受けて骨髄で産生され、その表面に発現しているL-セレクチン を介して血管内皮細胞に接着した後、ついで発現するPSGL-1と血管内皮細胞 のP-セレクチンが接着する。
骨髄のストローマ細胞もIL-1の刺激によりIL-5を産生することがわかっている。この時点での接着力は弱い(ローリング)。
なお、セレクチンにはもう一つE-セレクチンが知られているが、好酸球の遊走への関与はほとんどないと考えてよい。
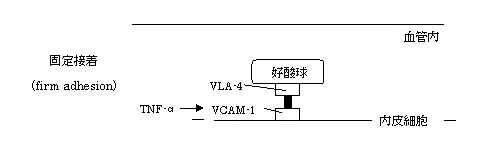
次に、好酸球のインテグリンVLA-4(α4β1)が細胞外基質であるVCAM-1を介して血管内皮細胞のインテグリンと接着する(固定接着)。VCAM-1の発現はTNF-αにより増強される。
β1インテグリンVLA-4(very late antigen-4)は接着のみならずVCAM-1やフィブロネクチンとの結合によりサイトカイン産生を介した好酸球生存延長を起こす。
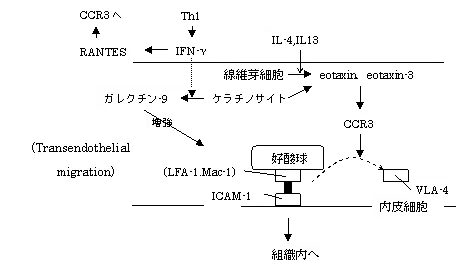
固定接着の後、繊維芽細胞やケラチノサイトが産生するケモカインeotaxin1,3らの作用で、好酸球表面にVLA-4に置き換わりLFA-1やMAC-1が発現する(α4の発現が低下し、β2の発現が増強される)。
eotaxin受容体はCCR3と一致する。また、eotaxinがCCR2に対するアンタゴニストで、CCR5に対するアゴニストである。
eotaxin2とeotaxin3はeotaxinと同様に好酸球の選択的遊走活性を有し、CCR3をそのレセプターに持つが、アミノ酸レベルでは40%以下程度の類似のみとされている。
eotaxin2は好酸球が血管内皮を移行する際の接着のシグナルがβ1インテグリンからβ2インテグリン優位にシフトするdetachmentに働く。
血管内皮細胞はIL-4の刺激によりeotaxin3のみをその表面に発現、産生し、抗CCR3抗体、抗eotaxin3抗体により好酸球の血管内皮遊走が阻害された。
LFA-1がICAM-1と接着することで好酸球は血管内皮細胞を通過する。
最近はα4β7とMAdCAM-1(expression of mucosal addressin cell adhesion molecule-1)の接着やPECAM-1(platelet endothelial cell adhesion molecule-1)の関与も指摘されている。
好酸球は、ICAM-1だけでなく、 β2インテグリン/ICAM-1を介して好酸球は単に接着するだけでなく活性化を受け脱顆粒、活性酸素産生、RANTES産生が惹起されること、GM-CSFを中心としたサイトカイン産生を介して好酸球生存を延長する。
また、β2インテグリンα鎖であるCD11b(CR3)の発現は、RANTES、PAF、IL-3、IL-5、GM-CSF、TNF-α、LPS、C5aなどにより増加、あるいは接着として機能的に増強すること、インテグリン相互のシグナル伝達はβ1インテグリンからβ2インテグリンへとなされていると考えられている。
ガレクチン-9はβガラクトシドに結合能を持つレクチンの一つで、好酸球遊走作用のほか、種々の免疫調節作用を持つ。 IFNγはは線維芽細胞からエカレクチン(ガレクチン-9)産生を誘導する。
好酸球の作用
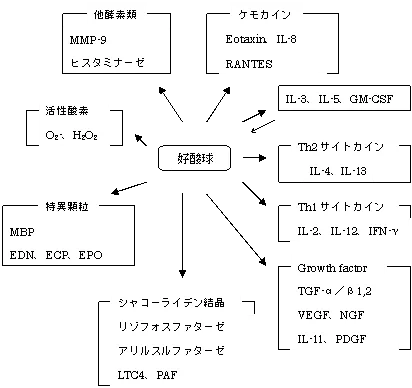
末梢血中から組織へと遊走した好酸球はIL-3、IL-5、GM-CSFという自分を活性化するサイトカインを産生したり、強塩基性タンパク質であるMBP(major basic protein)、EDN、ECP(eosinophil cationic protein)、EPO(eosinophil peroxidase)を特異顆粒から分泌する。特に特異顆粒のコアから分泌されるMBPの細胞傷害性は強力である。
なお、EDNは細胞傷害性は弱いものの、樹状細胞の分化(pre-DC→未熟DC→成熟DC)と遊走に深く関わっている。
<好酸球によるアレルギー・炎症反応の抑制>
アリルサルファターゼ・・・SRS-A、ECF-Aの不活化
ヒスタミナーゼ・・・ヒスタミンの不活化
ホスホリパーゼ・・・PAFをin vitroで不活化
EPO・・・LTの不活化
LTB4の白血球遊走活性をEPO+H2O2+halideで70%以上減少
catalase添加によりCaionophore刺激によるEoよりのLTの産生を増強
CLC・・・lysophospholipidを中和
PG・・・好塩基球からのヒスタミン遊離抑制
<好酸球によるアレルギー・炎症反応の促進>
EPO・・・EPO+H2O2+halideによる細胞傷害性、肥満細胞の分泌刺激作用
MBP・・・顆粒タンパクの53%を占める。crystalloid granuleのcore parasite killing、気道上皮細胞の剥離
ECP・・・顆粒タンパクの30%を占める。亜鉛を含むcrystalloid granuleのcore以外に存在。ヘパリンの不活化。
EDN・・granular matrix、Gordon現象
LT・・・LTC、気管過敏性亢進、遊走活性
PAF・・・気管過敏性亢進、遊走活性
好酸球の表面には、種々のレセプターが発現しており、好酸球は細胞障害性を発揮するために、これらのレセプターを修飾させている。
ヒト好酸球では、活性化した好酸球として”hypodense eosinophil(hypo Eo)と呼ばれるsubpopulationが存在する。 好酸球の脱顆粒において、顆粒タンパクを特異的に細胞外に放出する機構がこのhypo Eoの出現に関与している。 GM-CSFなどのサイトカインを加えることでhypo Eoを誘導することができる。
好酸球は免疫グロブリン受容体を有していて、IgG、IgE、IgAそれぞれの刺激で放出されるECP、EPO、EDNの脱顆粒に選択性がある。 さらに、分泌型IgAで刺激された好酸球はIL-4、IL-5、IL-10などのTh2サイトカインが特異的に放出され、CD28刺激ではTh1サイトカインのみが分泌される。
好酸球が抗原特異的・CD80/86依存性にCD4+細胞に抗原提示しうることも報告されている。
好酸球のレセプターは以下の通り
| レセプター | 内訳 |
| Fcレセプター | FcγR、FcαR、FcεR |
| 補体レセプター | C3a、C5a |
| インテグリン | Mac-1、LFA-1、VLA-4 |
| サイトカインレセプター | IL-3、IL-5、GM-CSF |
| ケモカインレセプター | CCR1、CCR3 |
| エイコサノイドレセプター | CysLT1、B-LTR、CRTH2 |
| ヒスタミンレセプター | H1、H2、H4 |
| その他 | PAFR、FAS、HLA-DR、fMLPなど |
好酸球の分泌顆粒、メディエーター、サイトカインは以下の通り
| 分泌物 | 内訳 |
| 活性酸素群 | OH-、O2-、H2O2 |
| 顆粒蛋白 | MBP、ECP、EDN、EPO |
| 脂質メディエーター | LTC4、LTB4、5-HETE、PGE1、PGE2、TxB2、PAF |
| サイトカイン ケモカイン |
IL-1,2,3,4,5,6,8,10,12,16,GM-CSF,TGF-α/β,RANTES,MIP-1α/β |
好酸球とアラキドン酸カスケード
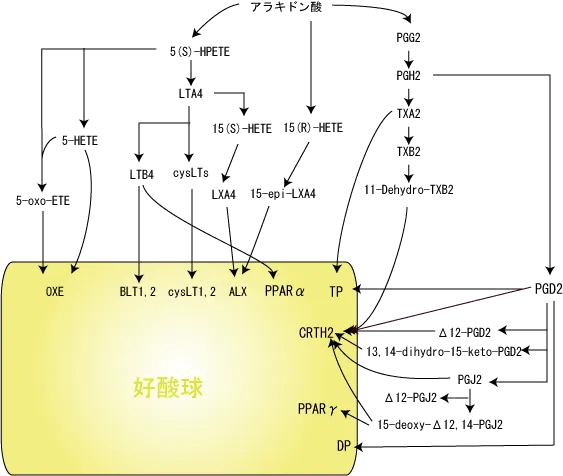
好酸球はLTC4などの脂質メディエーター産生細胞と共に、これらの脂質メディエーターにより直接あるいは間接的に自身の機能調節を受けている。 シクロオキシゲナーゼ産物のなかで最も好酸球との関連が強いのはPGD2である。
PGD2受容体DP(DP1)はGs蛋白と共役しているDPを介して、好酸球遊走抑制、生存延長などの作用を示すのに対して、CRTH2(DP2)を介したシグナルはGi蛋白と共役し、 好酸球遊走促進、CD11b発現増加、脱顆粒促進などの作用を示す。
DP1の炎症抑制作用は樹状細胞のDP1を介して制御性T細胞が誘導されるためとされている。
いずれにしても、DP1、CRTH2受容体の役割(DP1が抑制でDP2が活性など)は実験モデルにより結果が異なっており、不明な点が多いという。
DP受容体のリガンドはPGD2だけであるが、CRTH2受容体のリガンドはPGD2とその代謝物、TXA2やPGF2の代謝物など幅広く認識する。
LTB4は核内受容体であるPPAR(peroxisome proliferator-activated receptors)αを介して好酸球活性化を抑制する作用もある。
樹状細胞のDP1を介して制御性T細胞が誘導される。
PGD2の代謝産物の一つであるΔ12-プロスタグランジンJ2も好酸球動員作用を示す。
- CD
- 樹状細胞
- NK細胞
- リバウンド
- B細胞
- キラーT細胞
- 感作
- リンパ球
- アラキドン酸カスケード
- Igの構造
- サイトカイン
- ニューロペプチド
- セラミド
- 界面活性剤
- 好酸球
- ケモカイン
- インテグリン
- レギュレーターリスト
- 色素沈着
- 成長因子(GF)
- 活性酸素(ROS)
- NF-κB
- アレルギーの種類
- 骨代謝
- 抗菌ペプチド
- ヘルパーT細胞
- 好塩基球
- 薬の正しい使い方
- 血液凝固反応
- テオフィリン・β刺激薬の抗炎症作用
コメントor補足情報orご指摘あればをお願いします。
- << 前のページ
- 次のページ >>
