

キラーT細胞
体が外部抗原と戦う因子はマクロファージ、好中球、NK細胞、抗体と来て、後もうひとつキラーT細胞がある。
キラーT細胞はCD8+T細胞の別称であり、抗原提示を受ける前のナイーブT細胞である。
これはT細胞により呼び覚まされた後、自己の腫瘍細胞表面のクラスⅠ分子をキラーT細胞上のCD8で認識し、抗ウィルス作用を示す。(TCRと抗原、MHCクラスⅠとCD8の組み合わせ)
樹状細胞は単独でキラーT細胞に抗原提示できる(B7分子が常に発現しているため)が、その他の抗原提示細胞は、キラーT細胞がこのような細胞障害性(MHCクラスⅠを認識できる)を持つために、先にヘルパーT細胞がマクロファージ等の活性化によるB7分子発現まで終えておく必要がある。
ナイーブなキラーT細胞がエフェクターな細胞傷害性T細胞(CTL)になるためには、Th1が産生するIL-2等が必要となる(IL-2がナイーブCD8+T細胞の受容体にくっつくことが必要)。CTLはNK細胞と同じくIFN-γを産生してがんやウイルスを攻撃する。
通常抗原提示細胞とT細胞にはPD-L1と呼ばれるタンパク質が発現していて、抗原提示細胞のB7とT細胞のPD-L1、抗原提示細胞のPD-L1とT細胞のPD-1といった形で抑制的(免疫寛容)に働いている。本来抗原提示細胞のB7はT細胞上のCD28と結合し、共刺激のシグナルを伝達するが、CD28に競合する形でPD-L1が抑制をかける感じ。
CTLはPD-1を発現している、ということは、抗原提示細胞のPD-L1と結合して免疫を抑制することが可能。そして、がん細胞はPD-L1を表面に持つので、CTLによる破壊を免れることができる。そのため、抗PD-1作用のあるオブジーボ等の薬が抗がん剤として使用されている。
クラスⅠ分子は全ての自己細胞上及び抗原提示細胞上に発現しているが、キラーT細胞が認識するのはあくまで“クラスⅠ+抗原”であり、クラスⅠ単体では攻撃できない。
そして、“クラスⅠ+抗原”の“抗原”が自己の細胞であった場合は、外部抗原でなく自分を攻撃することとなるが、通常は胸腺での分化の過程でアポトーシスにより殺されているので大丈夫。
で、活性化したキラーT細胞(CTL)の抗ウィルス作用は大きく2つ、パーフォリン/グランザイム経路とFas/FasL経路に分けられる。
パーフォリン/グランザイム経路・・・・・パーフォリンにより細菌膜に穴を開ける
Fas/FasL経路・・・・・・・・・・・・・FasLがFas発現細胞にくっつくとアポトーシス
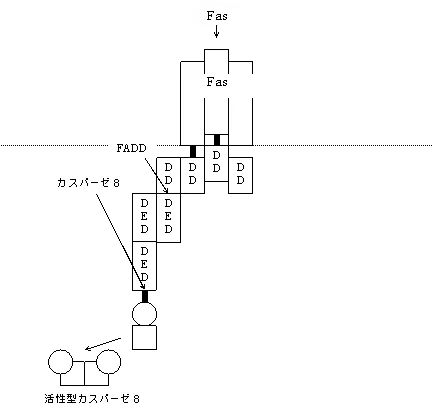
役目を終えたCTLは、他のエフェクターT細胞達とともに、大部分は死滅し、ごくわずかがエフェクターメモリーT細胞(TCM)とセントラルメモリーT細胞(TEM)に分化して、再度の抗原暴露に備える。
セントラルメモリーT細胞は、ナイーブT細胞ライクなT細胞で主に二次リンパ組織に常駐し、リンパ組織と血管間の移動に重要であったCCR7やCD62L(Lセレクチン)が発現している。
エフェクターメモリーT細胞は、主に末梢組織に常駐し、再度抗原暴露の際に素早く免疫反応を進行させる。遊走の必要がないのでCCR7やCD62Lの発現は低下している。
- CD
- 樹状細胞
- NK細胞
- リバウンド
- B細胞
- キラーT細胞
- 感作
- リンパ球
- アラキドン酸カスケード
- Igの構造
- サイトカイン
- ニューロペプチド
- セラミド
- 界面活性剤
- 好酸球
- ケモカイン
- インテグリン
- レギュレーターリスト
- 色素沈着
- 成長因子(GF)
- 活性酸素(ROS)
- NF-κB
- アレルギーの種類
- 骨代謝
- 抗菌ペプチド
- ヘルパーT細胞
- 好塩基球
- 薬の正しい使い方
- 血液凝固反応
- テオフィリン・β刺激薬の抗炎症作用
コメントor補足情報orご指摘あればをお願いします。
- << 前のページ
- 次のページ >>
