

抗原提示
マクロファージやB細胞、樹状細胞が抗原を貪食・分解した後、MHC分子と抗原ペプチドをT細胞へ受け渡すところです。
MHC分子
マクロファージやB細胞表面にあるMHC分子(人のMHC=HLA)には3つのサブタイプがありますが、抗原提示に関与しているのはクラスⅠとクラスⅡの二つでした。
MHCクラスⅠ
MHCクラスⅠ経路は、細胞内抗原(ウイルス・腫瘍由来タンパク)を、プロテアソーム→TAP→ER経路にて処理し、CD8陽性T細胞(キラーT細胞)に提示する経路です。
クラスⅠはほとんどすべての細胞に発現していますが、クラスⅡは限られた細胞にしか発現していません。
MHCクラスⅠ+抗原ペプチドは、キラーT細胞(CD8+)のCD8抗原により認識されるのだが、ヘルパーT細胞からの誘導なくしては活性化しない。
キラーT細胞はMHCクラスⅠ分子によって提示されるペプチド断片が自己であれば攻撃しませんが、非自己であれば細胞ごと破壊します。
各細胞内の不要なタンパク質がプロテアソームによりペプチド→アミノ酸まで分解されて、再び新しいタンパク質の生合成に利用されるサイクルとは別に、一部のペプチド(細胞構成タンパク質由来ペプチド)はアミノ酸まで分解されずにTAPトランスポーターと呼ばれる運搬分子によって小胞体に運び込まれる。
小胞体内で作られているMHCクラスⅠ分子は、小胞体にて先のペプチドを結合し、細胞表面まで運び、外部に向かって提示する。(これは、細胞内に存在するタンパク質をコードする遺伝子の塩基配列を知るのにも有用である。)
MHCクラスⅡ
MHCクラスⅡ経路は、外来抗原(細菌・ダニ抗原・異物)をエンドソーム経路により処理して提示する経路です。
MHCクラスⅡ分子はマクロファージや樹状細胞などの特定の細胞にのみ発現していて、ヘルパーT細胞(CD4+)に 対する抗原提示に関与している。
MHCクラスⅡ分子を持つ細胞達は、一般にエンドサイトーシスにより抗原を自細胞の中に取り込み、エンドソームと呼ばれる膜に囲まれた小胞の中でリソソーム酵素によってペプチドまで分解するが、ここで生成したペプチドがTAPトランスポーターを介して小胞体へ移動すると、これらの貪食細胞が破壊されてしまい、感染が拡大することになる。
そこで、クラスⅡによる抗原提示は、クラスⅠ経路のようなTAPトランスポーターによる輸送系には入らず、細胞外から取り込んだタンパク質由来のペプチドを含むリソソームとMHCクラスⅡ分子を運ぶ輸送小胞とが融合し、インバリアント鎖が外れることによって両者は結合→細胞表面まで運ばれる。
サイトカインによる自己活性化
マクロファージやDCが産生するサイトカイン等は自己やT細胞、B細胞を活性化する作用がある。
マクロファージの場合は、INF-α,β,γ、TNF-αらによりクラスⅠの発現が、INF-γによりクラスⅡの発現が上昇し、IL-10によりクラスⅡ発現が抑制される。
IL-10はTh1,Th2サイトカインを抑制、INF-γはTh2への分化を抑制(Th1とTh2のバランスの調整)。
B細胞の場合は、IL-4によってクラスⅡ発現が上昇し、INF-γにより抑制される。また、B細胞の分化を起こすためには、T細胞によるシグナル(IL-2)が必要である。
APCによる抗原提示
抗原提示先のT細胞には、
- CD4陽性T細胞・・・ヘルパーT細胞(Th1、Th2)
- CD8陽性T細胞・・・キラーT細胞
- CD4陽性T細胞亜群・・・Th17、NKT、Treg、Tr1他
があり、先に説明した通り、CD8+はMHCクラスⅠ、CD4+はMHCクラスⅡとなっている。
CD4+T細胞の場合、APCの細胞表面まで運ばれたMHCクラスⅡ分子はヘルパーT細胞のCD4抗原で、MHC分子と一緒に渡される抗原はT細胞表面にあるTCRで認識される。
つまり、CD4-MHCクラスⅡ、外部抗原-TCRの関係で認識される。
外部抗原-TCRにおける抗原は多種多様なため、TCR内の遺伝子はそのつど再構築される。
T細胞による抗原の認識にはもうひとつシグナルが必要で、それを共刺激シグナルと呼ぶ。
抗原提示細胞上のB7(CD80、86)分子とT細胞上にあるCD28分子 、ICAM-1とLFA-1、CD40とCD40L、VCAM-1とVLA-4などの抗原非特異的接着分子 が互いに認識しあうことで、抗原を提示できる(ハロー86にはニーハオ28で答える)。
まとめると、抗原を提示するときにT細胞と抗原提示細胞の間でやり取りされるシグナルは、
| <抗原提示細胞上> | <T細胞上> |
| MHCクラスⅡ | CD4 |
| 外部抗原 | TCR及びCD3 |
| B7(CD80、86)など | CD28など |

と言った具合。
CD8+T細胞であるキラーT細胞の方は、MHCクラスⅠとCD8、外部抗原とTCRの関係で認識される。
いずれにせよ、この3つのうちどれが欠けてもアナジー(不応答)を起こす。
TCRのシグナル伝達
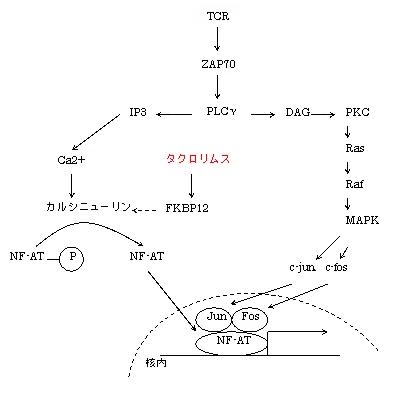
マクロファージ、樹状細胞らによる抗原提示が済むと、非受容体型PTKのLck活性化に引き続いて、同じく非受容体型PTKのZAP70が活性化します。
TCRはチロシンキナーゼ型受容体ではないので、2量体を形成して自身をリン酸化できません。そのため、TCRと複合体を形成しているCD3のITAMとよばれるチロシン残基を含むアミノ酸配列を他のアダプター分子等でリン酸化しなければなりません。
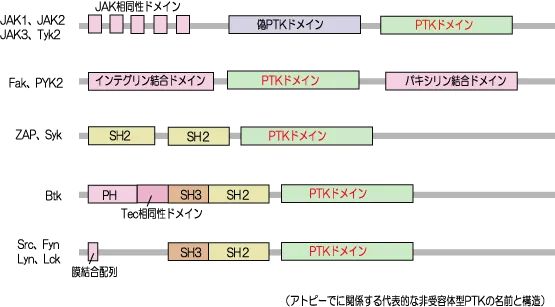
そこで、非受容体型PTKのSrcファミリーに属するLckが、チロシン残基のリン酸化を担い、そのリン酸基にSykファミリーに属するZAP70が結合し、Lckからリン酸化を受け活性化し、ZAP70はドッキングタンパク質のLATを介して、SH2ドメインを持つGrb2とPLCγを活性化します。
PLCγは自身のSH2で結合する以外にもPI3Kの下流でも活性化され、PIP2をIP3(イノシトール3リン酸)とDAG(ジアシルグリセロール)に分解することで、IP3は小胞体よりCa2+を遊離しカルシニューリンを活性化させ、カルシニューリンはNF-AT(nuclear factor of activated T cells)を脱リン酸化して活性化し、核内へ 移行して、がん原遺伝子であるc-fos、c-junを活性化し、これらをもとにタンパク質(Fos、Jun)が合成される。
がんタンパク質JunとFosはロイソンジッパーにより複合体(AP-1)を形成する。
活性化されたAP-1(activating protein-1:JunとFosの複合体)は、サイトカイン遺伝子のプロモーターに結合してIL-2等サイトカインの転写を促進する。
また、AP-1は、Aキナーゼに応答するcAMP応答配列(CRE)に結合する転写制御因子CREB(cAMP responsive element bind-ing protein)と結合して転写を促進するが、リン酸化されたCREBは、CREBの転写共役因子であるCBP(CREB-binding protein)を獲得し、さらに転写を促進する。
CREBの応答配列CREはAP-1結合部位の上流に位置している。
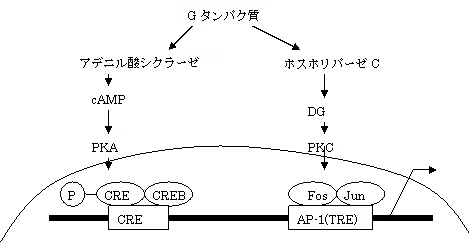
DAGはPKC(プロテインキナーゼC)を活性化して、PKCがRasやRaf-1を活性化することで、MAPKカスケードを進行させ、AP-1のリン酸化及び、NF-κBとIκB(inhibitor-κB)を解離させてNF-κBを遊離させる。NF-κBは核内に入りサイトカインを誘導する。
さらに、CREBはBcl-2の活性化によって、NF-κBはc-IAP1、Bcl-XLなどの遺伝子発現によりアポトーシスを抑制し、生存を促進する。
Grb2はSOS→Ras→Raf-1と進んでMAPKカスケードを進行させる。
これにより、マクロファージがさらに活性化するのは言うまでもないが、それ以外に、B細胞上のIg(イムノグロブリンのクラススイッチが起こることが非常に重要である。
TCRシグナル伝達のまとめ
TCRのシグナル伝達(花粉等アレルゲン→T細胞受容体(TCR)→サイトカイン合成→作用発現のルート)をまとめると以下。(こちらのサイトも詳しい)
- 1.アレルゲンを抗原提示細胞(マクロファージ、ランゲルハンス細胞、樹状細胞)がT細胞に受け渡す(アレルゲン-TCR、MHCクラスⅡ-CD4、CD86-CD28)
- 2.TCRと複合体形成のCD3分子のITAM(チロシン残基を2つ含むアミノ酸配列)が、非受容体型PTKのLckやFynによりリン酸化
- 3.そのリン酸基に非受容体型PTKのJAKやZAP70がSH2ドメインを介して結合
- PTK(protein tyrosine kinase)はチロシンキナーゼの略で、タンパク質のチロシンをリン酸化してシグナル伝達のON/OFFを切り替える酵素
- 受容体型チロシンキナーゼ:増殖因子(成長因子)の受容体。2量体を形成しお互いをリン酸化する。
- 非受容体型チロシンキナーゼ:代表はJAKやZAPといったPTKドメインを持つフリーのタンパク質で、サイトカイン受容体(JAK-STAT受容体)をリン酸化する。IL(インターロイキン)、造血因子などのサイトカインが結合し、それぞれのサイトカインの効能に関わる遺伝子の転写を促進する。
- 4.ZAP70はドッキングタンパク質のLATを活性化、LATはPLCγ(ホスホリパーゼCγ)やGrb2を活性化
- 5.PLCγはPIP2を、IP3とDAGに分解
- 6.IP3はCaストアからのCa放出することで、カルモジュリンの抑制が外れカルシニューリンが活性化、転写因子NFATが脱リン酸化による核内移行→NFATがDNA上のIL-2プロモーターに結合して、IL-2mRNAの産生が亢進する
- 7.DAGはPKC(プロテインキナーゼC)の活性化を介してRasやRafを活性化して、MAPKカスケードからのCREBの転写を促進するAP-1(Jun-Fos複合体)がリン酸化→核内へと移行しNFATやNF-κBらとともにサイトカイン(主としてIL-2)の合成を促進する。
- 8.Grb2はRasの活性化によりMAPKの活性化→⑦と同様の経路
- 9.作られたIL-2などのサイトカインはサイトカイン受容体に結合し、受容体とSTATが非受容体型PTKであるJAKによるリン酸化を受け、二量体を形成し、サイトカインに対応した転写因子を活性化する。JAK(ヤヌスキナーゼ)を阻害すればリン酸化からのサイトカイン合成が進まなくなる(ここがコレクチムの作用点)。
増殖因子のシグナル伝達(PI3K/Akt経路)については別ページ(チロシンキナーゼ型受容体)にて。
TCRからのシグナル伝達では非受容体型PTKが、PIP2をIP3とDAGに分解するところからのシグナル伝達に対して、増殖因子のシグナル伝達では受容体型PTKからの、PI3KがPIP2をリン酸化してからのシグナル伝達。
各サイトカインによる制御
T細胞にはCD4+のTh1とTh2、CD8+のTc以外にも、いくつかのCD4+T細胞亜群(Th3、Th17、Tr1、NKT、Treg)が存在する。サプレッサーT細胞は制御性T細胞(Treg)に取って代わっている。
もちらん、これらどのT細胞に抗原を提示するかによって最終的な生理作用は異なる。
Th2細胞は、
- IL-4、IL-13を出して抗体(免疫グロブリン=Ig)産生を誘導
- 肥満細胞の分化を促進
- IL-5を出して好酸球、好塩基球の分化を促進
- IL-6を出して血小板の分化を促進。IL-6はキラーT細胞を活性化するが、Th1細胞が産生するなんらかのサイトカインもキラーT細胞を活性化する。
Th1細胞は、IL-2、IFN-γを産生するとともに細胞性免疫を活性化する。

アレルギー疾患を考えるにはここも重要である。IgEへのクラススイッチの起こりやすさやTh2への傾きやすさなどの非MHC遺伝子によって、Th1よりTh2が優位になることでアレルギーが発現しやすい体質となるというのがひとつ。
また、T細胞に抗原提示する際、MHCが問題となることは先に述べたとおりであるが、人のMHC(HLA)には個体差(多型)があり、MHC型によって結合できるペプチド配列に偏りが生じる。ダニや花粉といった一部のアレルゲンに対してのみ免疫応答しやすいといった、アレルゲンとHLA型の相関のせいでアレルギーが発現しやすいというのもひとつである。
-
難易度低
├ 抗原vsマクロファージ
├ SOS
├ 第一次世界大戦?
├ MHCクラスⅡとTCR
├ T細胞の活性化
├ B細胞現る
├ 肥満細胞・好酸球・好塩基球
├ 皮膚をかくと?
└ 感作時
-
難易度高
├ 造血幹細胞
├ 接着
├ 非特異的免疫反応
├ 抗原提示
├ 肥満細胞とIgE
├ 補体の活性化
└ 細胞性免疫と体液性免疫
コメントor補足情報orご指摘あればをお願いします。
- << 前のページ
- 次のページ >>
