

DNAの複製(S期にて)
はれてS期へと進入すると、DNAが複製されます。ここで複製されたDNAが、M期での細胞分裂の際に2つの娘細胞に分配されます。
真核生物・原核生物共通で、動画にあるように、まず、DNAのらせん構造がⅡ型トポイソメラーゼによってほどかれます。

Ⅱ型トポイソメラーゼにはⅡA型(原核生物はDNAジャイレース、真核生物はtopoⅡ)とⅡB型(原核生物のみ)が存在しますが、どちらも下図のようにATPを加水分解してADPにする時のエネルギーを使ってDNAの2本鎖を切断してつなぎなおすことで、DNAのねじれを直していきます。

下表は細かいサブタイプ(wikiより)
| タイプ | サブタイプ | 原核生物(細菌) | 真核生物(人) |
|---|---|---|---|
| Ⅰ型 | IA 型 | topo I | - |
| topo III | topo III | ||
| (reverse gyrase) | - | ||
| IB 型 | - | topo I | |
| II 型 | IIA 型 | gyrase | - |
| topo IV | topo II | ||
| IIB 型 | (topo VI) | - |
ついで、Ⅰ型トポイソメラーゼの作用でDNAの1本鎖を切断してつなぎなおすことで、さらにDNAのねじれを直して複製しやすくします。Ⅰ型トポイソメラーゼにはⅠA型とⅠB型がありますが作用は同じです。


トポイソメラーゼⅠ型とⅡ型はどちらもDNAらせん構造のねじれを解消しますが、解消の仕方が異なるということを動画や図でチェックしておくとよいでしょう。
トポイソメラーゼⅡA型の内、原核生物のみに存在するDNAジャイレースを阻害する薬はニューキノロン系抗生物質です。DNAジャイレースは原核生物特有の酵素なので人には害を示しません(選択性)。
こうしてDNAの2本鎖はねじれがなくなり複製しやすい状態になるわけですが、複製を開始するためにはまだすることがあります。
それは、塩基同士の水素結合(下図の水色線)をDNAヘリカーゼ(大きく6つのファミリーがある)によって切断して、2本のDNA鎖の間を開くことです。開いた部分がフォークに見えることから、このY字の部分を複製フォークと呼びます。
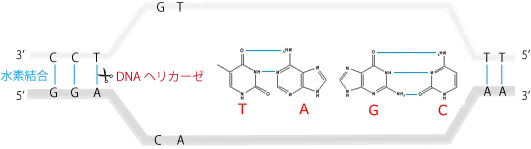
切断から複製までの過程は、真核生物、原核生物で異なるので別々に解説します。
真核生物のDNA複製
真核生物のDNAは直線状で、複製開始地点が複数あります。
しかし、複製開始の許可を与えるライセンスがないと複製が開始できません。
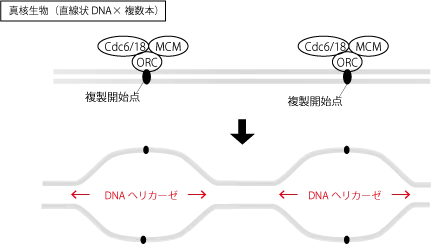
複製開始点にはORC(複製起点認識複合体:origin recognition complex)が結合していて、ここにCdc1/Cdc6タンパク質が結合し、その助けでMCM2-7ヘテロ6量体(minichromosome maintenance2-7)が結合します。
この3つからなる複合体をpreRC(複製前複合体:pre-replication complex)と呼びます。
preRCが活性化されると、DDK(Cdc7キナーゼ-Dbf4)が結合し、preRCのMCM2-7のN末端部分をリン酸化するとともに、CDKがSld2タンパク質とSld3タンパク質をリン酸化すると、Sld2/Sld3はDpb11と複合体を形成、Mcm10が呼びこまれ、それが引き金となってCdc45とGINSがpreRCに呼びこまれる。
Mcm2-7にCdc45とGINSが結合することで、Mcm2-7はDNAヘリカーゼ活性を持ち、2本鎖の水素結合を切断してDNAを1本鎖ずつに開裂させる。
その後、Cdc45がDNAポリメラーゼαとRNAプライマーゼを呼び込むことで複製がスタートする。(参考)
次にRNAプライマーゼによってRNAプライマーが作られます。
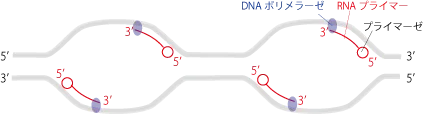
プライマーができるとDNAポリメラーゼがRNAプライマーを基にして、DNAを伸長します。
DNAポリメラーゼは何もないところからDNAを伸長できないので、最初2-10程度の塩基からなる短いRNAプライマーを一次的に作り、プライマーの3'末端のOHに新たなヌクレオチドをつなぐ形で伸長していき、最後にRNAプライマーを除去するという手順を踏みます。
ここで注意するのは、DNAの伸長は3'末端のOH側でなければならないため、DNAポリメラーゼが動く方向は必ず5'→3'方向でなくてはならないことです。
しかしながら、DNAヘリカーゼによって複製フォークが出来上がる方向は、上の鎖と下の鎖で5'方向と3'方向で異なるので、片方の鎖は開裂する方向とは逆方向へ複製していかなければならなくなります。
この時、開裂の方向を3'末端として伸長する上の鎖をリーディング鎖、開裂の方向が5'末端方向なので逆方向に伸長する下の鎖をラギング鎖といいます。
リーディング鎖は螺旋が開かれると同時に伸長(5’末端→3’末端方向へ)できるため、伸長スピードは速いです。
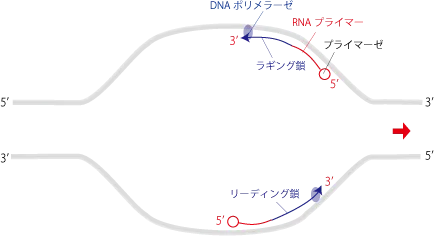
ラギング鎖の方は伸長方向が、3’→5’末端には伸ばせないので開裂方向とは逆方向へ伸長して行き、隣のRNAプライマーを飛び越えて隣のDNA断片まで到達し、DNAリガーゼによりフラグメント(岡崎フラグメント)をつないでラギング鎖を作ります。ひと手間が入る分、伸長スピードは遅いです。
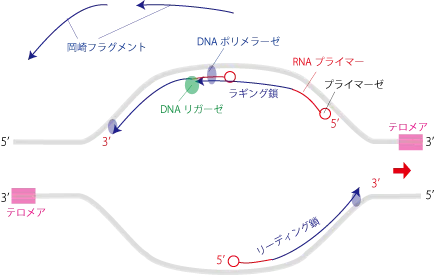
DNAの複製ではこのようにRNAプライマーを使用して伸長し、後からプライマーを取り除くため、新しくできる2本のDNAの5'末端側がRNAプライマー分だけ短く複製されます。
DNAの5'末端側にはTTAGGGの繰り返し配列であるテロメアと呼ばれる部位が存在し、この部分が細胞分裂のたびに短くなることからテロメアが寿命を表すと言われます。ただ、生殖細胞、幹細胞、がん細胞にはテロメラーゼと言われるテロメアを伸長することができる酵素があるため永遠に増殖することが可能です。
原核生物のDNA複製
基本的な伸長の過程は真核生物と同じですが、最初のDNAの螺旋構造を開く過程のみが異なります。
原核生物のDNAは輪ゴムのような環状構造であり、DNAの複製地点は一か所です。原核生物のDNAは染色体DNAとプラスミドDNAがありいずれも1つの環状DNAを持ちます。プラスミドDNAは全ての細菌が持っているわけではありません。

ORC(複製起点認識複合体:Origin recognition complex)と呼ばれる複製開始地点にDnaAタンパク質が結合し、ヘリカーゼ活性を持つDnaBタンパク質がDnaCタンパク質とともに、螺旋構造を開きます。
DnaBはヘリカーゼスーパーファミリー4に該当し、細菌のような環状DNAの複製に主に関わっている。ヘリカーゼファミリーは他も合わせて6つあり、このケースはそのうちの一つに過ぎない。
DNA複製阻害関連薬
- カンプト、トポテシン、オニバイド(イリノテカン)・・・抗悪性腫瘍薬。生体内でカルボキシルエステラーゼにより活性代謝物(SN-38)に加水分解されるプロドラッグである。Ⅰ型DNAトポイソメラーゼを阻害することによって、DNA合成を阻害する。
- ランダ(シスプラチン)・・・抗悪性腫瘍薬。癌細胞内のDNAと結合し、DNA合成及びそれに引き続く癌細胞の分裂を阻害するものと考えられている。DNA2本鎖の間に架橋を形成。
- マイトマイシン(マイトマイシンC)・・・抗悪性腫瘍薬。腫瘍細胞のDNAと結合し、二重鎖DNAへの架橋形成を介してDNAの複製を阻害し抗腫瘍効果を示すと考えられている。
- ゾビラックス(アシクロビル)・・・抗ヘルペスウイルス薬。ウイルス性チミジンキナーゼにより一リン酸化された後、細胞性キナーゼによりリン酸化され、アシクロビル三リン酸(ACV-TP)となる。ACV-TPは正常基質であるdGTPと競合してウイルスDNAポリメラーゼによりウイルスDNAの3’末端に取り込まれると、ウイルスDNA鎖の伸長を停止させ、ウイルスDNAの複製を阻害する
- バリキサ(ガンシクロビル)・・・抗サイトメガロウイルス薬。作用機序はアシクロビルと同じ。
- キロサイド、スタラシド(シタラビン:Ara-C)・・・抗悪性腫瘍薬。体内で活性代謝物のAra-Cに代謝された後、腫瘍細胞内に取り込まれ、デオキシシチジンキナーゼによりリン酸化されてAra-CMPになる。Ara-CMPは数回のリン酸化を受けて、Ara-CTPとなり、DNAポリメラーゼによりDNAに取り込まれることでDNA合成を阻害する。
シタラビン大量投与によりトランスポーターを介する膜透過が亢進し、細胞内シタラビン濃度が上昇し、薬剤耐性を克服できる。
- DNAと遺伝子とは
- DNAの転写
- タンパク質の合成
- 初期mRNAを作るまで
- チロシンキナーゼ型受容体
- JAK-STAT型受容体
- TGF受容体
- TNF受容体
- Wnt受容体
- Notch受容体
- Toll-like受容体
- NOD-like受容体
- RIG-like受容体
- MAPKカスケード
- タンパク質の分解
- 癌抑制遺伝子p53
- CKIとは?
- 細胞周期
- DNAの複製
コメントor補足情報orご指摘あればをお願いします。
- << 前のページ
- 次のページ >>
