

DNAと遺伝子とは
DNAとは別名デオキシリボ核酸と呼ばれ、真核生物においては核の中に存在します。原核生物では性染色体とプラスミドがこれに該当、ウイルスではコアの中に存在します。(細胞小器官)
核酸にはDNA以外にもRNAってのもあります。これはデオキシがつかないただのリボ核酸ですね。 それぞれの生物におけるDNAの存在場所がわかったところで、もう一度DNAとは何か考えてみる。
| 糖 | 塩基 | ヌクレオシド | ヌクレオチド (1リン酸) | リンの数 | ||
| デオキシリボース (DNA) | アデニン | Dアデノシン | Dアデニル酸 | dAMP | dADP | dATP |
| グアニン | Dグアノシン | Dグアニル酸 | dGMP | dGDP | dGTP | |
| シトシン | Dシチジン | Dシチジル酸 | dCMP | dCDP | dCTP | |
| チミン | チミジン | チミジル酸 | dTMP | dTDP | dTTP | |
| リボース (RNA) | アデニン | アデノシン | アデニル酸 | AMP | ADP | ATP |
| グアニン | グアノシン | グアニル酸 | GMP | GDP | GTP | |
| シトシン | シチジン | シチジル酸 | CMP | CDP | CTP | |
| ウラシル | ウリジン | ウリジル酸 | UMP | UDP | UTP | |
上の表を見ていただきたい。
糖には、リボースとデオキシリボースがあり、塩基にはアデニン、グアニン、シトシン、ウラシル、チミンの5種類があります。
ヌクレオシドは【糖+塩基】で、ヌクレオチドは【糖+塩基+リン酸】です(ヌクレオシドの5'位にリン酸基が結合)。
ヌクレオシドには、アデノシン、グアノシン、シチジン、ウラシル、チミジンがあり、ヌクレオチドには、AMP、GMP、CMP、UMP、TMP、ADP、ATP・・・らがあります。
ヌクレオチドという呼び名ではリン酸基の数を示すことができないので、リン酸基の数を示したい場合は、ヌクレオシド二リン酸やヌクレオシド三リン酸のように呼びます。
ヌクレオシド一リン酸には別名があり、呼び名は上の表参照(例:アデノシン一リン酸はアデニル酸(AMP)のこと。チミジン一リン酸はチミジル酸(dTMP)のこと)。cAMPは環状AMPのことでアデニル酸シクラーゼでATPから合成される。
糖がリボースからデオキシリボースに変わると、デオキシヌクレオシドやデオキシヌクレオチドと呼ばれ、接頭語にデオキシをつけて、略称にはdをつけて表します(例:dATP,dGTP,dCTP,dTTP,dUTP)。これらを総称してdNTPと呼びます。
RNAはヌクレオチドが連結した化合物で、DNAはデオキシヌクレオチドが連結して2本平行ならせん構造をとった化合物です。RNAの場合、塩基のチミンがウラシルに変わった点に注意です。
染色体とおなじじゃんと思うでしょうが、染色体は【DNA+ヒストン】であることをお忘れなく。
まとめると、ATPはヌクレオチドでありヌクレオシド三リン酸で糖がリボースで、dATPはデオキシヌクレオチドでありデオキシヌクレオシド三リン酸で糖がデオキシリボースという感じです。
リボースの構造はデオキシリボースのOH、HがOH、OHに変わっただけです。リボースはペントースリン酸経路から供給され、デオキシリボースはリボースの還元により得られます。
DNAの構造(二重螺旋構造)
さて、そんなDNAを図にしてみたのが下です。

糖とリン酸からなるバックボーンから4種類の塩基が手を出すように突き出していますね。DNAの塩基はA(アデニン)、G(グアニン)、C(シトシン)、T(チミン)の4種類があり、二重らせん構造の中でAとT、GとCがそれぞれ向かい合い、AとTは2本、GとCは3本の水素結合で結合しています。この水素結合を切るのはDNAヘリカーゼと言います。
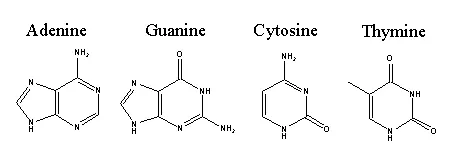
このDNAの螺旋構造をもっとミクロな視点から見てみると、まず塩基の化学構造はこんな感じです。
ここでよく見ると骨格のつくりが大きく2種類に分けられるはずです。アデニンとグアニンはプリン骨格を持ち、シトシンとチミンはピリミジン骨格を持ち、その骨格ごとに作られる経路が大幅に異なります。
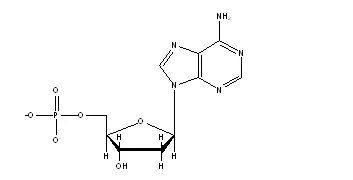
ついで、この塩基がリン酸と糖(デオキシリボース)とくっついたヌクレオチドはこんな感じです。
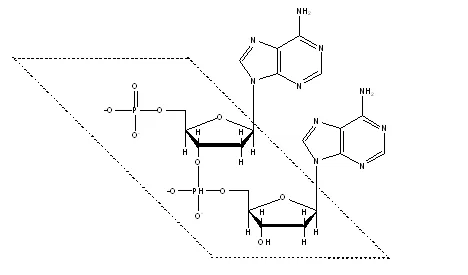
そして、このヌクレオチドが互いに結合して上のような2重螺旋構造をとったものをDNAと呼ぶわけです。点線でくくった部分が螺旋ということになります。図でいう左上のリン酸基が5'末端、左下デオキシリボースのOHが3'末端です。
DNAはOHがある3'末端側に伸長していくのですが、dNTPだけでは上のような構造(デオキシリボースのOHと燐酸が結合)がずーっと続いているだけで永遠に長くなってしまい染色体として完成されません。
しかし、dNTPとともにごく少量だけ存在している、デオキシリボースのOHがHに変わったものであるddNTP(ddATP,ddGTP,ddCTP,ddTTP)が結合すると、OHがないためにここで伸長がとまります。
ヌクレオチド代謝(de novo合成=生合成)については、以下の図を参考にしてください。
プリン合成
プリン骨格を持つ塩基(アデニン、グアニン)、キサンチン、尿酸の代謝経路。
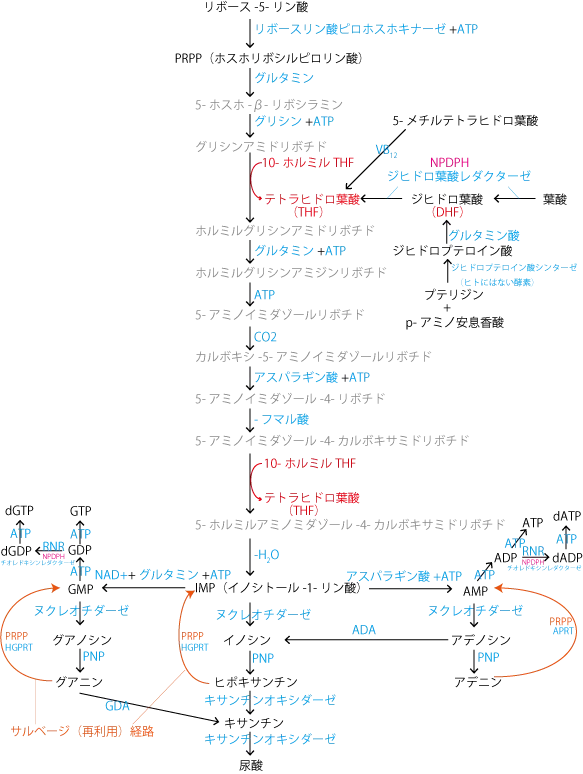
- プリン合成は、ペントースリン酸経路由来のリボース-5-リン酸に、ATPのピロリン酸基が転移して、PRPP(ホスホリボシルピロリン酸)を合成することからスタートする。
- PRPPはいくつかのアミノ酸(グルタミン、グリシン、アスパラギン酸)、CO2、10-ホルミルテトラヒドロ葉酸(10-ホルミルTHF)と多数のATPとGTPの作用により、IMP(イノシトール-1-リン酸)に変換される。
- この過程で使われる10-ホルミルTHFは、ホルミル基CHOをグリシンのアミノ基に転移(ホルミルグリシンアミドリボヌクレオチド→最終的にIMPへと変換)してテトラヒドロ葉酸(THF)になる。
THFは再びフォルミル化されて10-ホルミルTHFに再合成されてプリン代謝を回す。
THFはC1単位の転移反応(メチル基やホルミル基の転移)の補酵素として重要だが、実際に転移しているのは10-ホルミルTHFである。
THFはピリミジン代謝においても5,10-メチレンTHFを合成することでチミジル酸(dTMP)の合成に関わっている。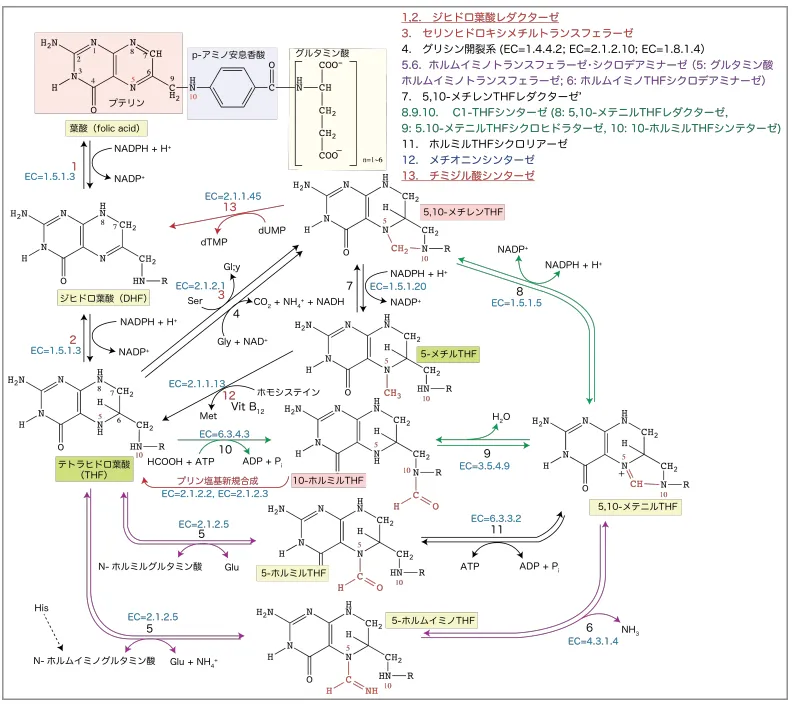
(ヌクレオシド代謝より引用)
細菌ではヒトにはないジヒドロプテロイン酸シンターゼ活性があるのでグアノシンから合成される2-アミノ-4-ヒドロキシ-6-ピロホスホリルメチルプテリジン(プテリジンの一つ)とp-アミノ安息香酸からTHFを生合成できる。(選択毒性)
葉酸はTHFを起点として様々な葉酸化合物が合成できるので、葉酸から合成しなくても、生体内で保存されている5-メチルTHFをビタミンB12とメチオニンシンターゼの作用で変換しても合成できる(この際ホモシステインがメチオニンに変換される。→メチオニンはSAMへ→クレアチニンの合成やグルコーゲン分解を促進する)。 - IMPからは、AMP、GMP、尿酸が作られる。
- AMP(アデノシン-1-リン酸)は、アスパラギン酸とATPの作用で合成される。AMPはヌクレオチダーゼによってリン酸基が加水分解されてアデノシンになる。アデノシンはPNP(プリンヌクレオシドホスホリラーゼ)で脱リボシル化されてアラニンになる。
- GMP(グアノシン-1-リン酸)は、NAD+とグルタミン、ATPの作用で合成される。GMPもヌクレオチダーゼによってグアノシンとなり、PNPによってグアニンになる。グアニンはGDA(グアニンデアミナーゼ)で脱アミノ化されてキサンチンになる。
- 尿酸は、IMPの5'リン酸が5'ヌクレオチダーゼによって加水分解され合成されるイノシンが、PNPによってヒポキサンチンになり、ヒポキサンチンがキサンチンオキシダーゼで酸化されてキサンチンに、キサンチンがさらにキサンチンオキシダーゼで酸化されると尿酸に代謝される。この反応は主に肝臓で行われる。
イノシンはIMPから直接合成されるのとは別経路で、アデノシンのADA(アデノシンデアミナーゼ)による加水分解によっても生成します。(アデノシン+H2O→イノシン+リン酸) - プリン骨格を一から作るエネルギーを節約するために、遊離塩基を再利用する経路のことをサルベージ経路という。具体的にはPRPPに、HGPRTやAPRTといった酵素の作用で遊離の塩基(グアニン・ヒポキサンチン・アデニン)を転移させてヌクレオチド(GMP、IMP、AMP)を再合成するということ。
- AMPとGMPがリン酸化されてできるリボヌクレオシド二リン酸(ADPとGDP)は、リボヌクレオチドレダクターゼ(RNR)の作用で還元されてデオキシリボヌクレオシド二リン酸(dADPとdGDP)になる。この反応は還元型チオレドキシンが触媒する。還元型チオレドキシンは酸化型チオレドキシンがNADPHを補酵素としてチオレドキシンレダクターゼの作用を受けて生成し、還元反応後は再び酸化型に戻る。
デオキシリボヌクレオシド三リン酸(dATPやdGTP)はデオキシリボヌクレオシド二リン酸のリン酸化で作られる。ATPやGTPから作られるわけではない点が重要である。
関連薬の作用点
- ロイケリン(メルカプトプリン)・・・抗悪性腫瘍薬で白血病の適応のみ。メルカプトプリン(6-MP)は細胞内に取り込まれた後、チオイノシン酸から6-チオグアニンヌクレオチド(6-TGN)に変換されると、イノシン酸(IMP)の構造類似体としてIMPの代わりに取り込まれ、GMPとAMPの合成を抑制する。また、チオイノシン酸及びそのメチル化体は、5-ホスホリボシル-1-ピロリン酸(PRPP)から5-ホスホリボシルアミンへの形成反応等プリンヌクレオチド合成に不可欠な反応を阻害する
- アザニン、イムラン(アザチオプリン)・・・免疫抑制薬で臓器移植、潰瘍性大腸炎、リウマチ、自己免疫性肝炎の適応。生体内で6-MPに分解され、核酸合成を阻害することにより免疫抑制作用をあらわす。
- メトトレキサートとトリメトプリムはDHFレダクターゼを阻害してTHFを減らすことで、10-ホルミルTHF等のC1転移反応の供与体の再合成を抑制→プリン代謝の抑制と、5.10-メチレンTHFの合成を抑制→チミジル酸(dTMP)合成(ピリミジン代謝)の抑制。
- アロプリノールはキサンチンオキシダーゼを阻害して尿酸の生成を抑制する。
- ブレディニン(ミゾリビン)・・・免疫抑制薬。プリン合成系のイノシン酸からグアニル酸に至る経路を拮抗阻害することにより核酸合成を抑制する41)が、高分子核酸中には取り込まれない
- セルセプト(ミコフェノール酸モフェチル)・・・免疫抑制薬。ミコフェノール酸モフェチル(MMF)は、生体内で速やかにミコフェノール酸(MPA)に加水分解される。MPAは、de novo系、salvage系2つのプリン生合成経路の内、de novo経路の律速酵素であるイノシンモノホスフェイト脱水素酵素を不競合的、可逆的かつ特異的に阻害することにより、GTP、デオキシGTPを枯渇させ、DNA合成を抑制する。T、Bリンパ球細胞は核酸合成を主としてde novo系に依存するのに対して、免疫系以外の細胞はde novo、salvage両系に依存している。MPAはsalvage系酵素には影響しないため、結果的にリンパ球細胞の増殖を選択的に抑制し、臓器移植後に発症する拒絶反応の形成不全を誘導する。
ピリミジン合成
ピリミジン骨格を持つ塩基(シトシン、チミン、ウラシル)の代謝経路。
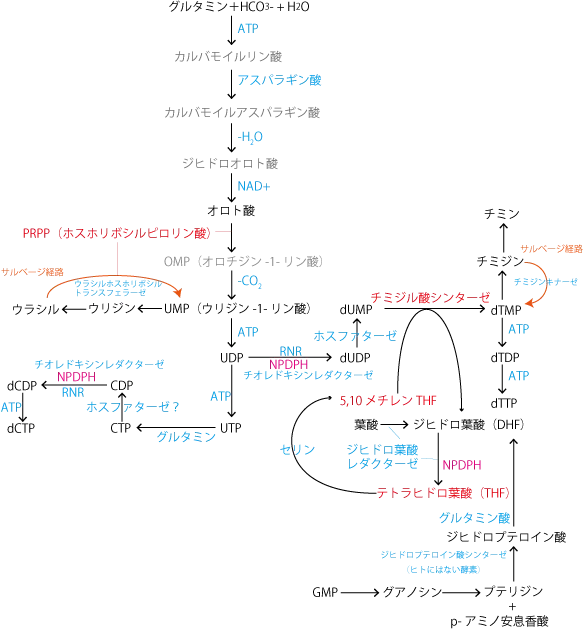
- ピリミジン合成はグルタミンとアスパラギン酸、炭酸イオンから、ピリミジン骨格を持つオロト酸を合成することからスタートする。
- オロト酸はプリン合成でも使われるリボース-5-リン酸由来のPRPP(ホスホリボシルピロリン酸)からホスホリボシル基を転移されたあと、脱炭酸反応を受けてUMP(ウリジン-1-リン酸)を生じる。
- UMPはATPによるリン酸化を受けて、UDP→UTPになり、UTPはグルタミンと反応してアミノ基がピリミジン環に結合し、CTP(シチジル-3-リン酸)が生じる。
- CTPが脱リン酸化されてCDPを生じたあと、プリン合成で述べたリボヌクレオシド二リン酸(ADPやGDP)の還元と同じ機構でデオキシリボヌクレオシド二リン酸(dCDP)となる。
- UDPについても同様にしてdUDPが作られた後、ホスファターゼで脱リン酸化されてdUMPになり、チミジル酸シンターゼの存在下、5,10メチレンTHFによるメチル基CH3の転移反応受けて、dTMPが作られる。この反応はウリジル酸からチミジル酸を作る反応として非常に重要。
- メチル基を転移した5,10メチレンTHFはジヒドロ葉酸(DHF)になり、ジヒドロ葉酸レダクターゼとNADPHによりTHFを再生し、セリンヒドロキシメチルトランスフェラーゼの作用でセリンをグリシンに変換する時にTHFに転移し、THFを5,10メチレンTHFに変換する。
- ピリミジン代謝でもサルベージ経路が存在する。一つ目はチミジンからチミジンキナーゼの作用でチミジル酸(dTMP)を再生する経路、もう一つはウラシルからUPRTによりウラジル酸(UMP)を再生する経路である。
関連薬の作用点
- フルオロウラシル(5-FU)・・・抗悪性腫瘍薬。代謝されて5-フルオロデオキシウリジル酸(5-フルオロdUMP)となり、チミジル酸(dTMP)の構造類似体として、チミジル酸シンターゼを阻害してdTMPの生成を抑制する。
- フルツロン(ドキシフルリジン)・・・抗悪性腫瘍薬で5-FUのプロドラッグ。腫瘍組織で高い活性を有する酵素、ピリミジンヌクレオシドホスホリラーゼ(PyNPase)により5-FUに変換され、抗腫瘍効果を発揮する。5-FUはFdUMPに代謝され、ウラシル由来のdUMPと拮抗し、チミジル酸合成酵素によるDNA合成経路を阻害する。また、5-FUはFUTPに変換され、ウラシルと同じくRNAにも取り込まれてF-RNAを生成し、RNAの機能を障害すると考えられている
- エスエーワン、ティーエスワン(テガフール・ギメラシル・オテラシルカリウム)・・・抗悪性腫瘍薬でテガフールが5-FUのプロドラッグで本体で他は作用増強剤。テガフール・ギメラシル・オテラシルカリウム配合剤(以下、配合剤)はFT、CDHP及びOxoの三成分を含有する製剤であり、経口投与後の抗腫瘍効果は体内でFTから徐々に変換される5-FUに基づいている。
CDHPは主として肝に多く分布する5-FU異化代謝酵素のDPDを選択的に拮抗阻害することによって、FTより派生する5-FU濃度を上昇させる。この生体内5-FU濃度の上昇に伴って、腫瘍内では5-FUのリン酸化代謝物である5-フルオロヌクレオチドが高濃度持続し、抗腫瘍効果が増強する。また、Oxoは経口投与により主として消化管組織に分布してorotate phosphoribosyltransferaseを選択的に拮抗阻害し、5-FUから5-フルオロヌクレオチドへの生成を選択的に抑制する。その結果配合剤投与により5-FUの強い抗腫瘍効果を損なうことなく消化器毒性が軽減されると考えられている。 - ゼローダ(カペシタビン)・・・抗悪性腫瘍薬。がん細胞でのみ活性体の5-FUに変換。消化管より未変化体のまま吸収され、肝臓でカルボキシルエステラーゼにより5’-DFCRに代謝され、その際に二酸化炭素が生成する。次に主として肝臓や腫瘍組織に存在するシチジンデアミナーゼにより5’-DFURに変換される。更に、腫瘍組織に高レベルで存在するチミジンホスホリラーゼにより活性体である5-FUに変換され抗腫瘍効果を発揮する。
XELOX療法(カペシタビン・オキサリプラチン療法) - キロサイド、スタラシド(シタラビン:Ara-C)・・・抗悪性腫瘍薬。体内で活性代謝物のAra-Cに代謝された後、腫瘍細胞内に取り込まれ、デオキシシチジンキナーゼによりリン酸化されてAra-CMPになる。Ara-CMPは数回のリン酸化を受けて、Ara-CTPとなり、DNAポリメラーゼによりDNAに取り込まれることでDNA合成を阻害する。
シタラビン大量投与によりトランスポーターを介する膜透過が亢進し、細胞内シタラビン濃度が上昇し、薬剤耐性を克服できる。 - サルファ剤(スルファメトキサゾール等)・・・p-アミノ安息香酸の構造類似体として細菌に取り込まれ、THFの生合成を阻害する。
遺伝子とは
DNAというものについては大体理解してもらえただろうか?人にしろ、動物にしろ、非常に長いDNA構造の中で実際に必要な部分は3割にも満たないといわれています。ここで言う実際に必要な部分を遺伝子といいます。
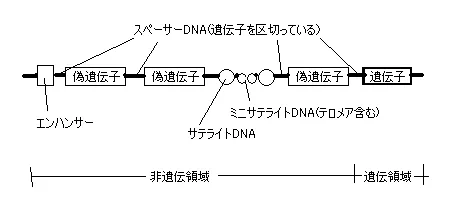
つまり、DNAの中にも多数の遺伝子(遺伝領域)といらない配列(非遺伝領域)があるということで、1生物すべての遺伝子のもつ遺伝情報を総称してゲノム(人ならヒトゲノム)と呼びます。
その構造を図式化したものが下図です。テロメアってのは細胞分裂するたび短くなっていくので、テロメアがなくなる=死ということになるらしいです。
そして、ある任意の遺伝子ひとつについて構造を図式化すると下のようになります。
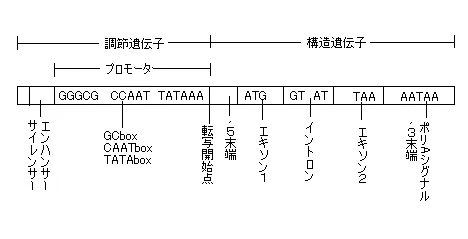
エキソン(タンパク質のアミノ酸配列に対する情報を持つ塩基配列)、イントロン(タンパク質のアミノ酸配列に関与しない塩基配列)は真核生物には共通な構造ですが、原核生物にはエキソンはありますがイントロンは存在しません。
- DNAと遺伝子とは
- DNAの転写
- タンパク質の合成
- 初期mRNAを作るまで
- チロシンキナーゼ型受容体
- JAK-STAT型受容体
- TGF受容体
- TNF受容体
- Wnt受容体
- Notch受容体
- Toll-like受容体
- NOD-like受容体
- RIG-like受容体
- MAPKカスケード
- タンパク質の分解
- 癌抑制遺伝子p53
- CKIとは?
- 細胞周期
- DNAの複製
コメントor補足情報orご指摘あればをお願いします。
- << 前のページ
- 次のページ >>
