

下行性抑制系
下行性抑制系にはセロトニン系とノルアドレナリン(NA)系の二つの系があります。
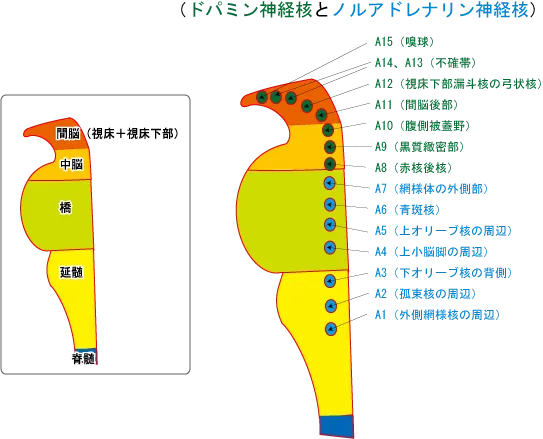
ノルアドレナリン系
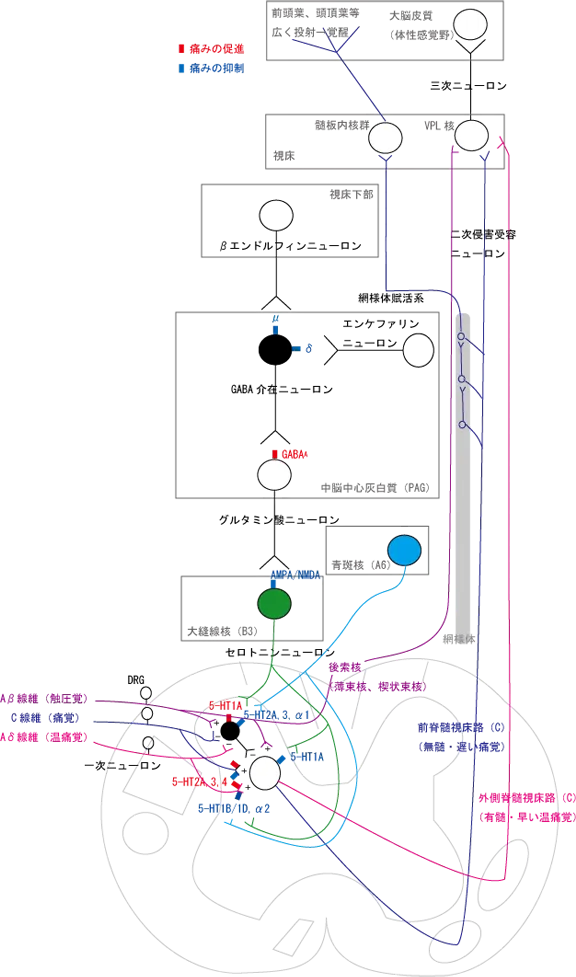
ノルアドレナリン系の方は、橋の外側にある青斑核から脊髄後角へ投射し、侵害受容ニューロンを抑制する。
脊髄後角のGABA介在ニューロンは、α1受容体を発現しており、青斑核ニューロンはここを賦活化させて二次侵害受容ニューロンを抑制することで鎮痛効果を発現する。
また、脊髄後角のAδ神経とC線維の神経終末にはα2受容体が発現しており、青斑核ニューロンはここをシナプス前抑制することで終末部からのグルタミン酸放出量を低下させて神経伝達を抑えることで鎮痛効果を発現する。
セロトニン系
セロトニン系のほうは単一の系ではなく、オピオイド(βエンドルフィン、エンケファリン、ダイノルフィン)、アミノ酸(アスパラギン酸、グルタミン酸)、GABA(γアミノ酪酸)ニューロンが絡んでくる。
視床下部の弓状核から投射されているβエンドルフィンニューロンは中脳周囲灰白質(PAG)のGABA作動性の抑制性介在ニューロンに投射→μ受容体を介して下行性抑制系の抑制を抑制(活性化)する。
βエンドルフィンの合成は、ストレスとPCMCの項で述べた通り、視床下部の室傍核からのCRHを起因としたPOMCのプロセッシングによって下垂体前葉から分泌される。
同様に、エンケファリンニューロンも、δ受容体を介して介在性ニューロンを抑制する。
抑制性の介在性ニューロンが抑制されることで、アミノ酸ニューロンが興奮して→延髄の大縫線核からのセロトニンニューロンを活性化させて、脊髄後角の下行性抑制系を賦活化させる。

痛みや触覚等の知覚神経は、主に非侵害性の知覚である触圧覚を伝えるAβ線維、侵害性の知覚のうち、イタッとかアツッのような早い温痛覚を伝える有髄のAδ線維、慢性疼痛の原因である遅い痛覚を伝える無髄のC線維に大別できる。
ゲートコントロール理論によれば、Aβ線維とAδ、C線維を同時に賦活させることはできず、片方の神経を活性化させれば、もう片方の神経伝達は抑制される。つまり、肌をさするようなAβ線維への入力は、抑制性の介在性ニューロンを活性化させることで、二次侵害受容ニューロンを抑制してC線維等による痛みの伝達を抑制する。
脊髄後角において、
- 二次侵害受容ニューロンに5-HT1Aが発現、ここを刺激すると疼痛が抑制される。
- 抑制性介在ニューロンに5-HT1A、5-HT2A、5-HT3が発現、5-HT1A刺激で疼痛促進、5-HT2A、5-HT3刺激で疼痛抑制される。
- 一次ニューロン終末に5-HT1B/1D、5-HT2A、5-HT3、5-HT4が発現、シナプス前抑制にて5-HT1B/1D刺激すると疼痛抑制、、5-HT2A、5-HT3、5-HT4刺激で疼痛促進される。
セロトニンの疼痛に関する役割は、疼痛促進系と疼痛抑制系が混在していているが、疼痛抑制系のほうが強いと言われる。縫線核セロトニン神経の細胞体に自己抑制性の5-HT1A、神経終末に放出抑制性の5-HT1B/1Dも発現していると思われるので、余計ぐちゃぐちゃになってくる。
一次ニューロンの神経細胞体は脊髄後根神経節(DRG)がその役割を担っていて、ここがあらゆる受容体の発現をコントロールしている。
一次ニューロンと二次ニューロンの伝達物質はDRGの細胞体により異なっていて、慢性疼痛を引き起こすC線維のDRGは小型細胞で、伝達物質としてグルタミン酸・SP、CGRPが使われる。Aδ線維やAβ線維はSPやCGRP反応性ではない非ペプチドニューロン(つまり、伝達物質はグルタミン酸のみ)と言われる。
グルタミン酸の受容体であるNMDA受容体は通常はMg2+によって抑制されているため、AMPA受容体からのNa+チャネルを介した神経伝達のみが行われていますが、SP等のペプチド類がNMDA受容体に結合していくとMg2+が外れてNMDA受容体が活性化し、Ca2+が流入→痛覚過敏を引き起こす。
リリカとメマリー
ここで、このCaチャネルのα2δサブユニットに結合し、Caの流入を抑制して、神経伝達物質の放出を抑えることで、一次ニューロンと二次ニューロンの伝達を遮断して、痛みを止める薬がリリカである。
一方、メマリーの方は末梢ではなく中枢(主として海馬付近)で働くのが少し異なります。脳内グルタミン酸受容体サブタイプのNMDA受容体チャネルの過剰な活性化を抑制することにより細胞内への過剰なカルシウムイオンの流入を抑制し、神経細胞傷害や記憶・学習障害を抑制すると考えられています。
二次侵害受容ニューロン
Aβ線維の一次ニューロンは脊髄後索から延髄の後索核(薄束核、楔状束核)とシナプスを形成して二次ニューロンへ接続後、視床(VPL核)へ、Aβ線維の一次ニューロンは脊髄後角で二次ニューロンへと接続した後、外側脊髄視床路を通って視床(VPL核)へ、C線維の一次ニューロンは脊髄後角で二次ニューロンへと接続した後、前脊髄視床路を通って視床(VPL核)へ投射され、三次ニューロン伝達されて大脳皮質(体性感覚野)へ投射され痛覚として処理される。
C線維は上行する際に中脳網様体部にも投射していて、そこから視床(髄板内核群)へ、汎性視床皮質投射により大脳皮質前野へ投射して覚醒を起こす(上行性網様体賦活系)。
-
神経路の理解
├ 神経の分類
├ 脳、辺縁系、基底核
├ 視床と視床下部
├ 中脳
├ 橋
├ 延髄
├ 運動路
├ 体性感覚路(体・四肢)
├ 体性感覚路(顔)
├ 視覚路
├ 聴覚路
├ 前庭感覚路
├ 上行性網様体賦活系
├ 下行性抑制系
└ 反射系
体性神経(運動・感覚神経)
├ かゆみの種類・・・中枢性のかゆみと末梢性のかゆみ
├ かゆみの伝達の概略
├ 神経線維の種類・・・かゆみを伝える神経とは
├ かゆみ受容器・・・かゆみ刺激を受け取る受容器
├ かゆみ受容体・・・TRPファミリーを中心に
├ 痒みと痛みの関係
├ 興奮伝導・・・かゆみの伝導メカニズム
├ DRGとガッセル神経節・・・神経節に存在するもの(NGFなど)
└ 脊髄後角と尾側亜核・・・一次ニューロンから二次ニューロンへ
自律神経(交感・副交感神経)
├ 自律神経というもの
├ 交感神経
└ 副交感神経
中枢神経(NA・Ad・DA・5HT・Ach・GABA)
├ ノルアドレナリン
├ アドレナリン
├ ドパミン
├ セロトニン
├ アセチルコリン
└ γアミノ酪酸
コメントor補足情報orご指摘あればをお願いします。
- << 前のページ
- 次のページ >>
