

セロトニン神経系
セロトニンの合成と分布
消化管粘膜(腸クロム親和性細胞)に約90%、血小板に約8%、中枢神経系に約2%。
セロトニンの産生
トリプトファンからトリプトファン水酸化酵素により、5-HTP(5-水酸化トリプトファン)へ、5-水酸化トリプトファン脱炭酸酵素によりセロトニンへ。
神経終末から分泌されたセロトニンは、一部は神経終末のセロトニントランスポーターから再吸収され、残り(約90%以上)がMAOA(モノアミン酸化酵素A)により速やかに分解されて、5-HIAA(5-水酸化インドール酢酸)へと代謝され、尿から排泄される。
脳内で水酸化酵素を持つのは、縫線核群の5HT神経か松果体のメラトニン産生細胞だけ。
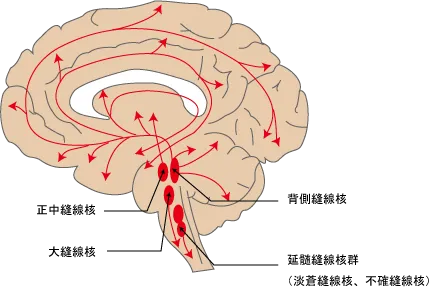
セロトニンの神経核
セロトニンの神経核は脳幹部にある縫線核群であり、ここにセロトニンの細胞体が存在し、各部位へとニューロンを伸ばしている。
縫線核群の分布とその番号と名称が以下。

| 神経集団番号 | 神経集団名称 | 主な作用 |
|---|---|---|
| B1 | 淡蒼縫線核 | 延髄縫線核群として、脊髄前角運動ニューロンへ(抗重力筋の増強)、脊髄中間外側核※1の交感神経節前ニューロンへ(交感神経興奮) |
| B2 | 不確縫線核 | |
| B3 | 大縫線核 | 脊髄後角へ(下行性抑制による鎮痛) |
| B4 | 舌下神経前位核の背側 | |
| B5 | 橋縫線核 | |
| B6、B8 | 正中縫線核 | 海馬に投射→記憶情報処理 |
| B7 | 背側縫線核 | 上行性に投射、大脳皮質(覚醒)、側坐核(衝動的行動)、前脳基底部(覚醒)、視床下部核群(睡眠、体温調節、摂食、内分泌)に投射 |
| B9 | 線状核 |
※1 胸髄の前角後側方の側角に交感神経節前ニューロンの細胞体の集合である中間外側核がある。
セロトニンの分泌要因
セロトニンの分泌亢進因子は、歩行、咀嚼、呼吸のリズム運動と太陽光である。
光は網膜から縫線核へ直接の軸索投射で、5HT神経を賦活する。(※視交叉上核のサーカディアンリズムとは異なる)
セロトニンの分泌抑制因子は、慢性ストレスである。ストレスによって活性化される神経である室傍核CRH神経が縫線核に投射していて、5-HT神経の活動を抑制する。ただし、一過性の覚醒・ストレス刺激に対しては影響されない。
セロトニンの作用
セロトニンは受容体だけでもサブタイプを含めて14種類あるため、その作用は非常に多様・複雑、そしてわかりづらいです。セロトニンはどんな物質?とは一言で言えないのが辛いところ。
覚醒・鎮静(抗不安)・睡眠の調節
| 関連受容体 | 5HT2A、5HT2C、5HT1A |
| 関連薬 | |
|---|---|
| セディール | 5HT1A刺激。シナプス後膜の5HT1A刺激による抗不安作用→抗鬱 |
| デジレル | SARI。5HT2A遮断、5HT再取り込み阻害、α遮断。5HT2A遮断→鎮静、5HT1A刺激→抗不安作用→抗鬱 |
| パキシル | SSRI。5HT再取り込み阻害。5HT1A刺激→抗不安作用→抗鬱 |
| トレドミン | SNRI。5HT,NA再取り込み阻害。5HT1A刺激、NA増加作用→抗不安作用→抗鬱 |
| トフラニール | 三環系。5HT,NA再取り込み阻害、α遮断、抗コリン、H1遮断。5HT1A刺激、NA増加作用→抗不安作用→抗鬱。心毒性あり。効果発現はSSRIよりも遅く、約1ヶ月以上はかかる。 |
| テトラミド | 四環系。α2遮断、α1遮断、弱い抗コリン。α2遮断作用によるNA放出促進→抗鬱。α1遮断→セロトニン拮抗。 |
| ルジオミール | 四環系。NA再取り込み阻害(5HTは再取り込み阻害作用なし)。NA濃度上昇→抗鬱 |
| リフレックス | NaSSA。5HT2、5HT3遮断、α2遮断。α2遮断→NA・5HT放出促進→抗鬱。5HT2、5HT3遮断→5HT1A選択性亢進。 |
| ジプレキサ | MARTA。D2遮断、5-HT2A,2B,2C、5-HT6遮断、α1遮断、H1遮断、弱い抗コリン、弱いD1遮断、弱い5HT3遮断。5HT2A/2C遮断によるD1活性化による抗鬱、抗不安。5HT2A遮断が睡眠障害改善、H1遮断が鎮静作用、5HT2C遮断とH1遮断が食欲減退の改善。 |
※SSRI等再取り込み阻害薬により、シナプス間隙のセロトニンが増加するとシナプス前膜の5HT1Aと5HT1B/1Dのオートレセプターを介したネガティブ・フィードバックが働くが、反復投与による脱感作(1~2週間)で抑制が外れると、セロトニンの分泌が上昇し、シナプス後膜の5HT1Aを介した抗不安効果が発現する。
シナプス後膜の5HT2受容体のダウンレギュレーションと抗鬱効果についてはまだ明らかではない。
抗鬱効果を発揮するためにはシナプス後膜の5HT1A受容体の活性化が重要。5HT1A受容体の刺激作用は、抗うつ作用・抗不安作用を示すとともに、前頭前野のドパミン放出を促進し、認知・陰性症状を改善すると考えられている。
背側縫線核のセロトニンニューロンは持続的な自発インパルスを発する等、覚醒レベルを調節している。
細胞体に存在している5HT1A受容体は自己抑制レセプターとして機能し、セロトニンの過剰の分泌を抑制する作用がある(もちろん、シナプス後膜の方にも5-HT1A受容体は存在している)ため、ここを刺激するセディールのような薬剤は、覚醒を抑制する。
運動不足、引きこもりのようなセロトニン分泌不足や慢性ストレスによる分泌抑制によってセロトニン欠乏脳が続くと、5HT1A受容体数が増加するが、そのため、余計に5HT分泌能が低下する。実際、うつ病患者の5HT1A受容体は増加している。
他視交叉上核に投射しているので、メラトニンの分泌調節(覚醒)と交感神経興奮あたりが起こると思われる。5HT2A、5HT2C遮断作用は睡眠障害を改善する。
生物時計の同調(覚醒と睡眠)
| 関連受容体 | 5HT1B、5HT7 |
視交叉上核の5HT1B受容体は網膜視床下部路からの入力で生物時計の同調。サーカディアンリズムには5HT1Bが関与する。
また、行動性同調機構として摂食行動、運動も視床・外側膝状体から視交叉上核に投射する経路で5HT7を介して作用。(視交叉上核の5HT7受容体)も関与。
視交叉上核からの命令で、松果体にて作られる睡眠ホルモン(メラトニン)は、必須アミノ酸のトリプトファンからセロトニン→N-メチルセロトニンを経て体内で合成される。メラトニンの合成は光により抑制されている。
ロゼレムはメラトニン受容体(MT1及びMT2)受容体を刺激して睡眠を誘発する薬。
レム睡眠とノンレム睡眠
| 関連受容体 | 5HT1A、5-HT1B、5-HT2A/2C、5-HT3、5-HT6、5-HT7 |
セロトニンは覚醒時には規則的な発射があるが、徐波睡眠(ノンレム睡眠時)に移行するとその活動が減弱、レム睡眠時には消失する。すなわちREM睡眠を抑制する。(参考:脳波)
セロトニンの神経とノルアドレナリン神経の活動停止が、中脳橋領域の、橋脚被蓋核(Ch5)、背側外側被蓋核(Ch6)のアセチルコリン神経の持続的な活動を可能として、それがREM睡眠状態を一定時間発現する。
前脳基底部のマイネルト基底核(Ch4)へのセロトニン投与は、δ波を優位にしてノンレム睡眠を発現させる。
背側縫線核の5HT1Aによる自己抑制は、中脳腹側被蓋野のコリン作動性神経を脱抑制(興奮)させて、レム睡眠を発現する。
セロトニンは、アセチルコリン神経(背側経路と腹側経路の両方)を抑制し、背側経路抑制によるレム睡眠の誘発と、腹側経路抑制によるノンレム睡眠の発現→レム睡眠への移行を導くということになる。
睡眠改善作用
| 関連受容体 | 5HT2A |
5HT2A受容体を遮断すると、入眠潜時の短縮(寝付きの良さ)と入眠効率の改善に働く。
| 関連薬 | |
|---|---|
| リフレックス | Nassa。5HT2A/2C、3遮断作用、α2遮断作用、H1遮断作用 |
睡眠時無呼吸
| 関連受容体 | 5HT2 |
5HT2は筋の緊張を維持して気道閉塞(無呼吸)を抑える。
衝動行動の抑制
| 関連受容体 | 5HT2A |
| 関連薬 | |
|---|---|
| リスパダール | SDA。D2、D3遮断、5HT2遮断。5HT2A、2Cを遮断してD2遮断薬の副作用の錐体外路障害を改善し、D3を遮断して帯状回のD2活性化→意欲回復、D1を賦活して認知機能改善。少量で弱い抗鬱(適応外) |
腹外側前頭前野(外側前頭前野の腹側部)に5HT2Aが存在する。5HT2Aを刺激すると衝動攻撃行動を抑制(ドパミン抑制)できるが、欠乏するとキレやすくなる(ドパミン活性)。これはセロトニン神経がドパミン神経を抑制していることに起因する。
うつ病脳では長期間5HT分泌不足による、5HT2A受容体の数が増加している。
5HT2Aは、ドパミン前頭葉のD1や基底核に向かうD2受容体を抑制。→遮断すると前頭葉のD1活性化による認知障害と陰性症状(情意、自発、動作障害)を改善する。また、基底核に向かうD2遮断による運動副作用の緩和。そして、睡眠覚醒レベルを司るセロトニンを抑制して眠気を誘発する。
5HT2Aは、ドパミンによる快情動の誘発、陽性症状(幻覚、妄想)も抑制する。腹側被蓋野→側坐核のDA分泌に対しての抑制。
※統合失調症の陽性症状とは、ないことをあるとすること(過剰な考え)、陰性症状とは、あることがないこと(意欲の喪失)。
下行性抑制
| 関連受容体 | 5HT1A/1B/1D、5HT2A、5HT3、5HT4 |
| 関連薬 | |
|---|---|
| トレドミン | SNRI。5HT,NA再取り込み阻害。大縫線核(B3)からのセロトニンニューロン、青斑核(A6)からのNAニューロンの賦活化による疼痛抑制。 |
下行性抑制による鎮痛を参照
抗重力筋の促通(増強)効果
| 関連受容体 | 5HT |
5HTは抗重力筋を補強することで、姿勢を正す。抗重力筋とは重力に負けないように直立に必要な筋肉群のこと。
副交感神経から交感神経への切り替え
| 関連受容体 | 5HT |
延髄縫線核群の5HT神経から脊髄中間外側核の交感神経節前ニューロンに興奮性の入力がある。
記憶の抑制
| 関連受容体 | 5HT1A、5HT2A、5HT3、5HT4、5HT6 |
海馬には多様な受容体(5HT1A、5HT2A、5HT3、5HT4、5HT6)があり、記憶の選択と強化の際に海馬のθ波(脳波参照)が重要。θ波は通常の覚醒時には抑制されている。注意行動時に発現して記憶情報処理を促進、過剰な記憶の蓄積を防ぐため。
5HTは海馬GABA作動神経の5HT3受容体を刺激し、θ波を抑制している。
θ波が現れる原因は、正中縫線核からのセロトニン神経が抑制される状況(何かに注意を向けた時、新しいものを見ている時、興味をいだいた時)などに記憶過程が活性化される。
記憶の長期増強(LTP)の効率はθリズムのタイミングで与えると最も効率が良い。
5HT1Aは自己受容体刺激で記憶改善、シナプス後膜受容体刺激で空間学習効率悪化、5HT4は記憶改善、5HT6は刺激で空間学習効率悪化。
前脳基底部(マイネルト基底核)の5HT投与はα波の増加(通常の覚醒ではβ波、睡眠はδ波優位)。アルツハイマーにおける変性部位がここにあるので、障害されるとα波の周波数が減少する。
体温調節
| 関連受容体 | 5HT1A |
背側縫線核のセロトニンニューロンは体温調節中枢のある視床下部に投射している。
一般にセロトニン神経の亢進によって体温上昇が起こる(セロトニン症候群)が、オートレセプターである5HT1A受容体を刺激すると体温が低下する。
摂食調節(食欲抑制)
| 関連受容体 | 5HT1B、5HT2B/2C、H1 |
背側縫線核のセロトニンニューロンは摂食中枢(外側核)、満腹中枢(腹内側核)のある視床下部に投射している。
セロトニン神経は摂食行動を抑制している。5HT1B、5HT2C共に弓状核のPOMCニューロンの活動を活性化させることで摂食行動を抑制的に制御している。
視床下部の5HT2C受容体遮断(抑制の抑制)で食欲減退を改善する。またH1受容体遮断も食欲減退を改善する。5HT1BはPOMCの抑制ニューロンを抑制する間接作用。
5HT2C受容体遮断で、体重、食事量、血糖値が増加する。→視床下部の満腹中枢へ神経伝達が行かないことも原因。
胃の5HT2B/2C受容体を遮断するとグレリンの分泌が増加し、食欲が増進する。六君子湯は脳内のグレリン受容体を増加したり、抗癌剤によるグレリンの分泌阻害作用を阻害して食欲を増進させる。
運動機能調節
| 関連受容体 | 5HT |
背側縫線核のセロトニンニューロンは小脳への広範な投射により運動失調を改善する。
片頭痛の改善
| 関連受容体 | 5HT1D、5HT1B、5HT1F、5HT2A、5HT7 |
| 関連薬 | |
|---|---|
| イミグラン | 5HT1D/1B受容体刺激による血管収縮作用 |
| エルゴタミン | 5HT刺激による血管収縮作用 |
セロトニンは血管を収縮させるため、血管収縮→反跳性の血管拡張により引き起こされる片頭痛発作を治療する。
5HT1D、5HT1F(三叉神経)、5HT1B(軟膜血管平滑筋)を刺激すると脳血管が収縮、またCGRPの放出抑制による神経原性炎症抑制機序にて片頭痛が収まる。5HT2Aは血管平滑筋を収縮するものの炎症性痛覚を悪化させるので刺激は好ましくない。
5HT2、5HT7は血管拡張作用にて、片頭痛発作の開始に関与する。
嘔吐・腹痛・下痢
| 関連受容体 | 5HT3 |
| 関連薬 | |
|---|---|
| カイトリル | CTZ及び胃の求心性迷走神経終末等の5HT3受容体を遮断して嘔吐を抑制する。 |
| イリボー | 腸管の迷走神経神経節(遠心性)の5HT3受容体を遮断して胃腸運動を抑制して下痢を抑え、腸管の迷走神経終末(求心性)の5HT3受容体を遮断して脊髄・延髄への痛みの求心路をたち、下痢による腹痛や痛覚過敏を抑える。 |
| プリンペラン | 胃のD2受容体遮断作用、5HT3受容体遮断作用、5HT4刺激作用を併せ持つ。胃の求心性神経終末、副交感神経節やCTZの5HT3遮断作用により嘔吐反応を抑制、5HT4の消化管運動亢進作用とD2遮断によるAch↑。 |
EC細胞(腸クロム親和性細胞)より供給されたセロトニンは、知覚神経終末の5HT3受容体を介してマイスナー神経叢の求心性の迷走神経細胞体を刺激し、アウエルバッハ神経叢の介在ニューロン→運動ニューロンへと伝わり、胃腸運動を調節するとともに、延髄・脊髄へ投射して、遠心性の自律神経を介して再びアウエルバッハ神経叢へ投射され、胃腸運動を調節する。
嘔吐中枢は延髄網様体に存在していて、求心性の迷走神経の入力を受けると、孤束核、迷走神経背側運動核、疑核等に出力して遠心性迷走神経による胃の逆蠕動運動や腹圧の上昇等の嘔吐反射を引き起こす。嘔吐中枢→視床VPL核→大脳皮質へと入力されて、嘔気を引き起こす。
消化管運動の亢進
| 関連受容体 | 5HT4 |
| 関連薬 | |
|---|---|
| ガスモチン | 5HT4刺激作用による消化管運動亢進作用 |
迷走神経末端シナプス前膜上のD2受容体はアセチルコリンの放出を抑制する。逆にシナプス前膜上の5HT4受容体はアセチルコリンの放出を促進して消化管運動を亢進させる。

血小板凝集作用
| 関連受容体 | 5HT2A |
| 関連薬 | |
|---|---|
| アンプラーグ | 5HT2A受容体を遮断することで血小板の凝集を抑制する。 |
血小板の5HT2A受容体を刺激すると血小板が凝集する。引き金となるのはPAF等の物質。
- 参考・引用元
-
神経路の理解
├ 神経の分類
├ 脳、辺縁系、基底核
├ 視床と視床下部
├ 中脳
├ 橋
├ 延髄
├ 運動路
├ 体性感覚路(体・四肢)
├ 体性感覚路(顔)
├ 視覚路
├ 聴覚路
├ 前庭感覚路
├ 上行性網様体賦活系
├ 下行性抑制系
└ 反射系
体性神経(運動・感覚神経)
├ かゆみの種類・・・中枢性のかゆみと末梢性のかゆみ
├ かゆみの伝達の概略
├ 神経線維の種類・・・かゆみを伝える神経とは
├ かゆみ受容器・・・かゆみ刺激を受け取る受容器
├ かゆみ受容体・・・TRPファミリーを中心に
├ 痒みと痛みの関係
├ 興奮伝導・・・かゆみの伝導メカニズム
├ DRGとガッセル神経節・・・神経節に存在するもの(NGFなど)
└ 脊髄後角と尾側亜核・・・一次ニューロンから二次ニューロンへ
自律神経(交感・副交感神経)
├ 自律神経というもの
├ 交感神経
└ 副交感神経
中枢神経(NA・Ad・DA・5HT・Ach・GABA)
├ ノルアドレナリン
├ アドレナリン
├ ドパミン
├ セロトニン
├ アセチルコリン
└ γアミノ酪酸
コメントor補足情報orご指摘あればをお願いします。
- << 前のページ
- 次のページ >>
