

目次
- 脂肪酸と中性脂肪
- コメント
脂肪酸と中性脂肪
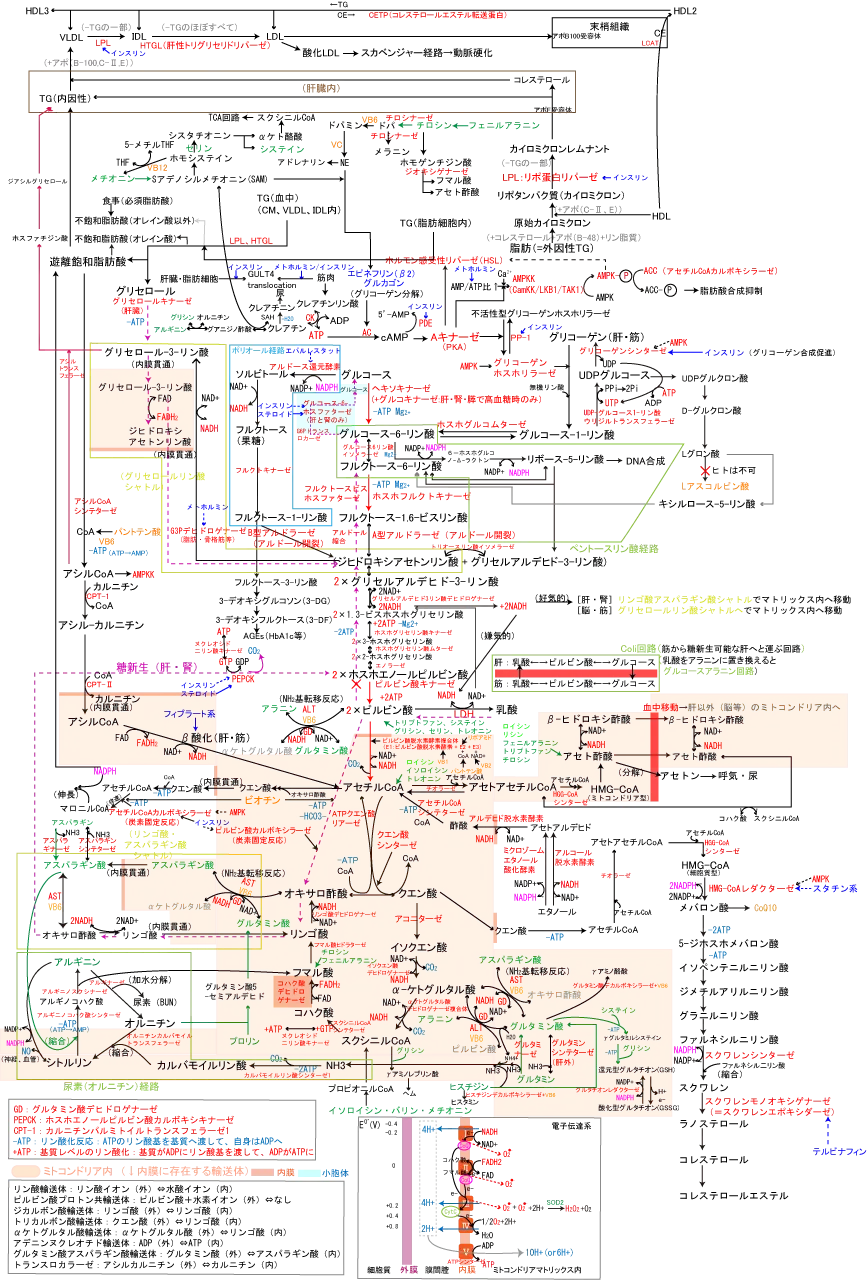
代謝全体マップは以下図参照。
脂肪酸
脂肪酸は上記中性脂肪やリン脂質の構成成分のひとつであり、炭素-炭素二重結合(>C=C<)の有無によって飽和脂肪酸(二重結合なし)と不飽和脂肪酸(二重結合あり)に分類される。
飽和脂肪酸(パルミチン酸(C16)、ステアリン酸(C18)等)は動物性である乳製品らに多く含まれ、炭素数が増えるほど融点が高いので、炭素が4つの酪酸(バターに含まれる)などは常温では液体ですが、炭素18個のステアリン酸は常温で固体です。飽和脂肪酸は体内で合成できるため、無理に取る必要はない。
一方、不飽和脂肪酸は植物油中に多く含まれ、二重結合を持ちます。
二重結合にはさまれたメチレン基(CH2)が酸化を受けやすいために体内で過酸化脂質になる(腐りやすい)こともしばしばである。動物体内で合成されない不飽和脂肪酸(n-3系列とn-6系列)を必須脂肪酸と呼び、リノール酸、α-リノレン酸がそれに該当します。
これらは体内で合成できないため、食事から摂る必要があり、摂取された不飽和脂肪酸は、体内で、
- n-6系(ω=6):リノール酸 → γリノレン酸 → ジホモγリノレン酸 → アラキドン酸
- n-3系(ω=3):α-リノレン酸 → EPA(エイコサペンタエン酸) → DHA(ドコサヘキサエン酸)
という風に代謝をされる(n-?は炭素-炭素二重結合の場所)。 不飽和脂肪酸の代表としては、オレイン酸(C18、2重結合=1)、リノール酸(C18、2重結合=2)、α-リノレン酸(C18、2重結合=3)がある。
脂肪酸の合成
脂肪酸の原料となるのは過剰な糖質やアミノ酸から生成するアセチルCoAです。脂肪酸の合成は細胞質で行われる。
ミトコンドリアマトリックス内で生成したアセチルCoAは、クエン酸シンターゼの作用によりオキサロ酢酸と縮合してクエン酸とCoAに変えられた後、クエン酸の形で内膜を通過し、細胞質でATPクエン酸リアーゼとATPの作用によりCoAと縮合してアセチルCoAとオキサロ酢酸に戻される。
こうして細胞質へ移動したアセチルCoAは、アセチルCoAカルボキシラーゼの作用でATPを消費してビオチンからカルボキシ基を受け取り、マロニルCoAを合成する。
ここで出来たマロニルCoAとアセチルCoA、NADPHを基質として、アセチルCoAをつないで(炭素鎖2ずつ延長)長鎖脂肪酸を合成する。(8アセチルCoA+14NADPH+7ATP→パルミチン酸+14NADP++8CoA+6H2O+7ADP+7Pi)。
C16より長い脂肪酸はC16をもとにしてミトコンドリアで進む。不飽和脂肪酸はヒトではC18(ステアリン酸)からオレイン酸(C18)のみ合成可能。リノール酸とαリノレン酸は生体内で合成できないので食事から摂取するしかない(必須脂肪酸)
合成された長鎖脂肪酸はアルブミンと結合して血中を移動し、脂肪細胞にTGとして保存される。
脂肪酸のβ酸化
β酸化は脂肪酸を分解してアセチルCoAを取り出すことである。脂肪酸のβ酸化はミトコンドリア内で行われる。
パルミチン酸等の遊離脂肪酸(FFA)をATPを使って(ATP→ADPでなくATP→AMPなので実質2ATP使用)アシルCoAにした後、ミトコンドリア内膜を通過できないアシルCoAをカルニチンでアシルカルニチンにすることで通過させ、マトリックス内で再度アシルCoAに戻す。
アシルCoAから大量のアセチルCoAを作るβ酸化の流れは以下である。
- アシルCoAデヒドロゲナーゼによってアシルCoAのα(2番目の炭素)とβ(3番目の炭素)の炭素間にあるシングルボンドを二重結合に変換する。この過程でFADがFADH2に還元され、トランス型の二重結合を持つエノイルCoAが生成する。
- エノイルCoAに水が添加されることで、3-ヒドロキシアシルCoAが生成される。
- 3-ヒドロキシアシルCoAはさらに変換されて、NAD+がNADHに還元される過程で3-ケトアシルCoAを生成する。
- 最後に、3-ケトアシルCoAは3ヒドロキシアシルCoAチオラーゼによってアセチルCoAと、もとの脂肪酸鎖から2炭素短縮された新しいアシルCoAが生成される。
パルミチン酸(C16)では7回のβ酸化が起こり、炭素2のアセチルCoAが8つとNADHとFADH2が7つずつ生成するため、NADHは2.5molのATP、FADH2は1.5molのATP換算、1molのアセチルCoAはクエン酸回路で10molのATPを生み出し、実質ATP消費が2molなので、合計106molのATPが生み出されることになる。
これはあくまで肝臓でのβ酸化で生じたアセチルCoAをそのまま肝臓でTCA回路に入れてATPを生成する場合であり、このアセチルCoAを肝以外の組織へエネルギー源として移動させるために水溶性で血中移動可能なケトン体(アセト酢酸、βヒドロキシ酢酸、アセトン)を合成する。
アセト酢酸はアセチルCoA2つが縮合したアセトアセチルCoA、さらに1つ縮合したHMG-CoAを経て作られ、βヒドロキシ酢酸とは可逆的に変換できともに血中移動可能だが、アセトンはエネルギー源とはならずに呼気や尿として排泄されるだけである。また、ここのHMG-CoAはミトコンドリア型であり、細胞質で行われるコレステロール合成の時の中間体のものとは型が異なる。肝臓で作られるケトン体は肝臓ではエネルギー産生つまりTCA回路には入れないので、糖新生はできないため、主に骨格筋へ移動してアセチルCoAに戻された後にTCA回路に入ってATPを産生する。
トリアシルグリセロール(TG=中性脂肪)
中性脂肪はトリグリセリド、トリグリセロール、トリアシルグリセロール、脂肪などと色々な別名でも呼ばれ、脂肪酸とグリセロール(グリセリン)から構成される。
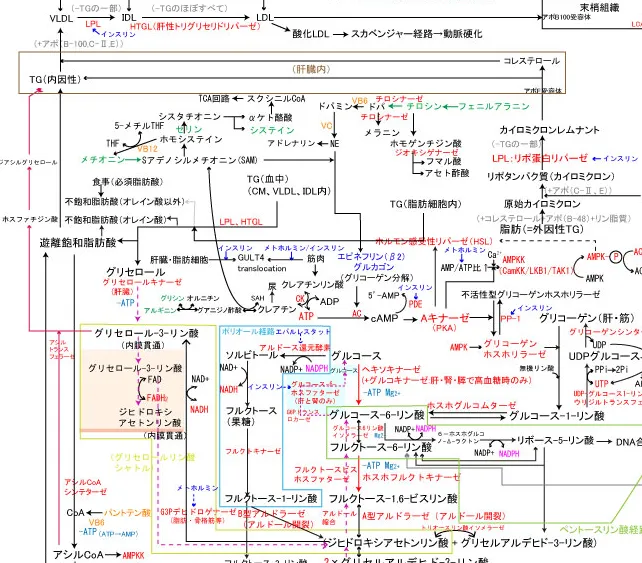
体内の中性脂肪は内因性のものと外因性のものに分けられ、内因性のものは、血中の過剰な脂肪酸が組織に取り込まれたり、肝臓で内臓脂肪となったりしたもので、血中リポ蛋白内のものもこれに属す。
外因性のものは、食事由来で、小腸で吸収されてから腸管で再合成され、肝臓を通らずリンパ管から血液中に出て行くものを指す。
結論から言えば、脂肪を減らすためには解糖系からの電子伝達系で賄いきれないエネルギー消費が行われる必要があるため、運動が必須。糖質が不足して脂肪のβ酸化が亢進してアセチルCoAが出来、ケトン体として骨格筋などへ移動してそこでエネルギーとして消費される。骨格筋で電子伝達系を回すには酸素が必要なため有酸素の運動が必要。糖質不足に関しては乳酸や脂肪分解で出てくるグリセロールなどから糖新生することで維持する形。
トリグリセリドの分解
トリグリセリドの分解は低血糖時に糖新生やβ酸化で必要なFFAやグリセロールを供給する反応として重要である。
トリグリセリドの分解は、リパーゼによりなされる。
具体的には、
- 膵リパーゼ:膵臓から分泌され、小腸での食事由来のトリグリセリドの分解。トリグリセリドを遊離脂肪酸(フリーファッティーアシッド(FFA))と2モノアシルグリセロールに分解する。
- リポプロテインリパーゼ (LPL):主に毛細血管の内皮細胞上で活性化されます。
この酵素は、キレミクロンやVLDLなどのリポプロテイン粒子に含まれるトリグリセリドを分解する役割を持っています。 LPLの活性化により、トリグリセリドはFFAとグリセロールに分解され、これらは周囲の組織(特に筋肉や脂肪組織)に取り込まれます。 - ホルモン感受性リパーゼ (HSL):HSLは細胞内のトリグリセリドの分解に関与しています。
この酵素は主に脂肪細胞内のトリグリセリドを目的としており、HSLの活性化により、トリグリセリドはFFAとグリセロールに分解されます。
グルカゴンやアドレナリンといったホルモンの影響でHSLは活性化され、トリグリセリドの分解が刺激されます。
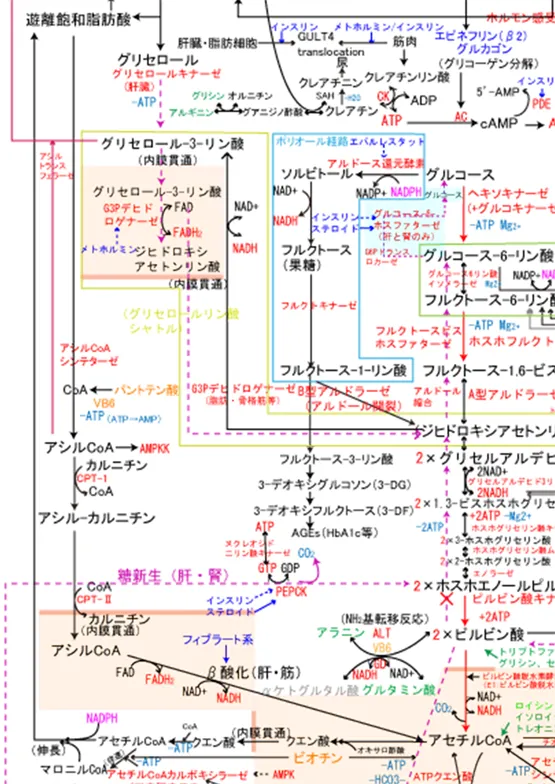
インスリンはPDE(ホスホジエステラーゼ)を活性化してcAMPをAMPに変えるのでHSLによる脂肪分解を抑制する。
メトホルミンは複合体を阻害することでATPを低下させ、AMP/ATPの比を高め、AMP活性化プロテインキナーゼ(AMPK)のγサブユニットがそれを検知し、αサブユニットのスレオニン172番残基(T172)を露出させ、T172がAMPKK(LXB1、CAMKK2、TAK1の3つ)によってリン酸化されるとAMPKが活性化され、AMPKはHSLを抑制的に制御し脂肪酸の動員を抑制したり、ACCを抑制し、アセチルCoA+マロニルCoAによる脂肪酸合成を抑制する。結果、血中遊離脂肪酸を減少させる。 - ヘパチン酸トリグリセリドリパーゼ (HTGL):HTGLは主にHDLやIDL内のトリグリセリドの分解に関与しており肝臓で発現している。
この酵素の活性により、リポプロテインのトリグリセリドがFFAとグリセロールに分解されることで、リポプロテインの成熟や変換が助けられます。
トリグリセリドの合成
トリグリセリドの合成は単純にグリセロールに3つの脂肪酸をくっつけることで行われるわけではない。
グリセロールは肝臓ではグリセロールキナーゼ活性があるのでグリセロール3-リン酸に直接変換できるが、脂肪細胞内にはグリセロールキナーゼが存在しないので、グルコースから作られるジヒドロキシアセトンリン酸をNADHで還元してグリセロール3-リン酸に変換される。
血中のFFAはアルブミンと結合する形で末梢の脂肪細胞へと移動して、アシルCoAシンテターゼで一旦アシルCoAに変えられた後にアシルトランスフェラーゼの作用でグリセロール3-リン酸に転移される。
脂肪細胞には褐色脂肪細胞と白色脂肪細胞の2つがあり、褐色脂肪細胞はミトコンドリアを持ち脇の下等特定の部位にのみ存在しATPを熱産生に使用するが、白色脂肪細胞はミトコンドリアをほとんど持たず、細胞内にTGをため込むだけである。
白色脂肪細胞内にはグリセロールキナーゼが存在しないのでグルコースからグリセロール-3-リン酸を作ってTGを合成する。脂肪細胞はインスリンによってGULT4が小胞体からトランスロケーションするのでグルコースを取り込めるようになり、結果TGが蓄積する。
グリセロール3-リン酸にアシルCoAのアシル基が1つ転移するとリゾホスファチジン酸が、さらにアシルCoAのアシル基を1つ転移してホスファチジン酸が作られ、ホスファチジン酸はホスファターゼでリン酸基を外されてジアシルグリセロールへ、ジアシルグリセロールはさらにアシルCoAからアシル基を受け取りトリアシルグリセロールが出来上がる。
参考サイト:薬学部学生向け講義資料ダウンロード
- 糖質の構造と種類
- 脂質の種類
- タンパク質の種類
-
ビタミン
├ 脂溶性ビタミン
| ├ ビタミンA
| ├ ビタミンD
| ├ ビタミンE
| ├ ビタミンF
| └ ビタミンK
|
└ 水溶性ビタミン
├ ビタミンB1
├ ビタミンB2
├ ビタミンB6
├ ビタミンB12
├ ビタミンC
├ ビタミンM
├ ビタミンH
├ ビタミンP
├ ビタミンQ
├ ニコチン酸
└ パントテン酸
コメントor補足情報orご指摘あればをお願いします。
- << 前のページ
- 次のページ >>
