

糖新生
糖代謝全体マップは以下図参照。
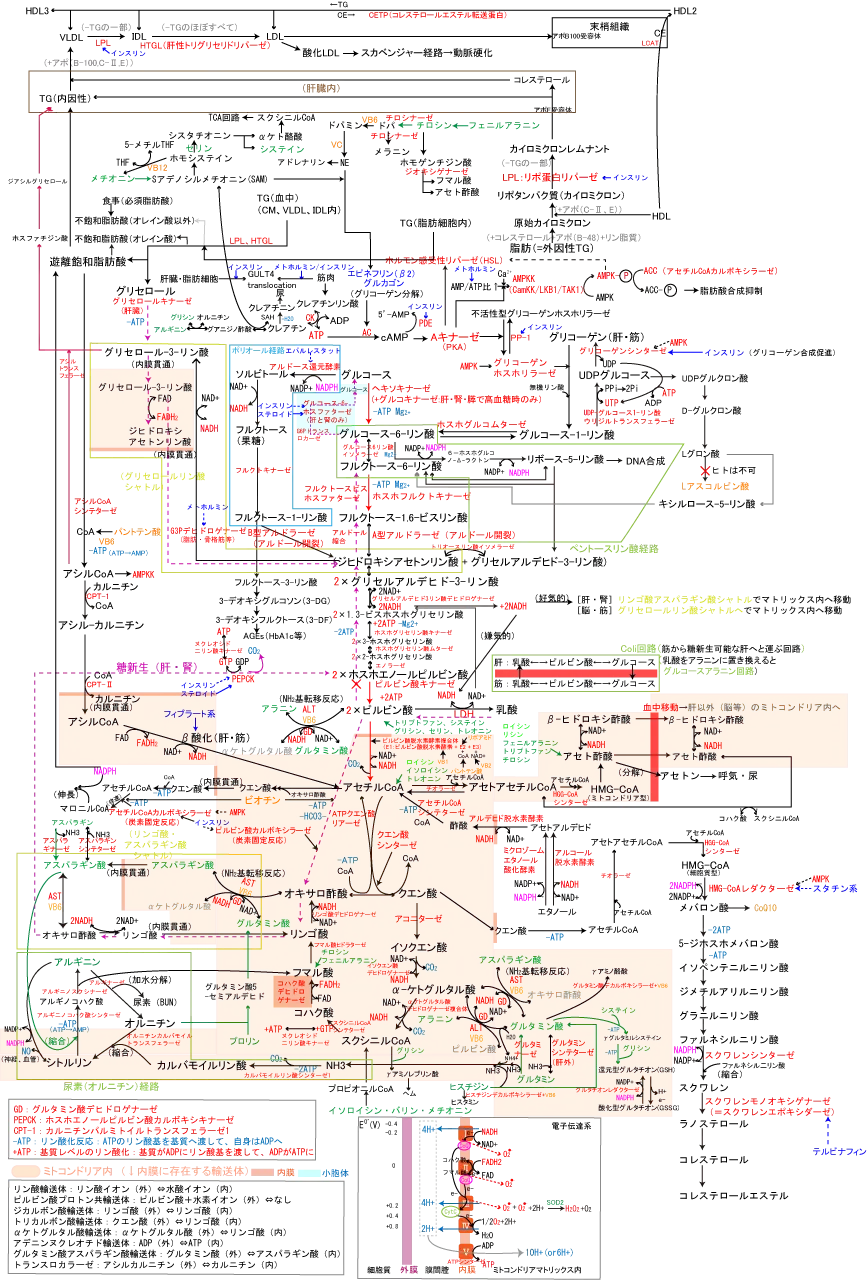
糖新生は糖以外の物質からグルコースを作る経路のことで肝臓と腎臓でのみ行われる。
糖新生の目的はグルコースという他の組織に移動可能なエネルギーを獲得することにある。
そのため、糖新生の間は肝臓のTCA回路は回っていない。つまり、肝臓でのエネルギー産生は行われていない。なぜならTCA回路を回すことを犠牲にしてグルコースを生み出すからであり、オキサロ酢酸を作るのと引き換えにαケトグルタル酸を消費するアミノ基転移反応はその代表である。
何もグルコースまで戻さずに、乳酸やケトン体、糖原生アミノ酸の形で移動してTCA回路に入ってエネルギーを取り出すことも可能だが、高強度の運動で酸素を利用しにくい時や、グルコースしか利用できない赤血球や大量にエネルギーを必要とする脳(脳はケトン体も利用できる)等で、グルコースは重要なエネルギー源として利用される。
糖新生のエネルギー源及び経路は、
- グリセロール→ジヒドロキシアセトンリン酸→グルコース
- ※グリコーゲン→グルコース1-リン酸→グルコース6-リン酸→グルコース
- 乳酸→ピルビン酸→グルコース
- 糖原生アミノ酸(アラニン、トリプトファン、システイン、グリシン、セリン、トレオニン)→ピルビン酸→グルコース
- 糖原生アミノ酸(グルタミン酸、グルタミン、アルギニン、ヒスチジン、プロリン)→αケトグルタル酸→リンゴ酸→グルコース
- 糖原生アミノ酸(イソロイシン、バリン、メチオニン)→スクシニルCoA→リンゴ酸→グルコース
- 糖原生アミノ酸(チロシン、フェニルアラニン)→フマル酸→リンゴ酸→グルコース
- 糖原生アミノ酸(アスパラギン酸、アスパラギン)→オキサロ酢酸→リンゴ酸→グルコース
が知られている(※グリコーゲン分解は正確には糖新生ではない)。
糖新生の共通の経路
糖新生が肝臓と腎臓でのみは行われる理由は、ホスホエノールピルビン酸からの解糖系の逆経路で最後のグルコース6-リン酸からグルコースを生成されるために必要な酵素(グルコース6-ホスファターゼ)が肝臓と腎臓にしか存在しないためである。
糖新生では、ピルビン酸からホスホエノールピルビン酸に直接戻れないことを理由にオキサロ酢酸経由で迂回してホスホエノールピルビン酸に戻った後、解糖系を逆に進行し、最終的にグルコースに戻される。
糖新生のスタートがどれであれ、この迂回から始まる解糖系の逆経路のどこかで合流することになる。
以下ピルビン酸からの逆経路
- ピルビン酸は、アセチルCoAかATPの濃度が上昇すると、TCA回路へは入らず、ピルビン酸カルボキシラーゼの作用でオキサロ酢酸へと変換される。この反応は炭素固定反応と呼ばれ、ATPを消費するとともに、補酵素としてビオチンが使用される。
- オキサロ酢酸がミトコンドリア内膜を通過して細胞質へ移動できないため、リンゴ酸デヒドロゲナーゼとNADHで還元されてリンゴ酸になる。リンゴ酸は内膜を通って細胞質へ移動し、NAD+で酸化されてオキサロ酢酸にもどされる
- 細胞質へ移動したオキサロ酢酸はホスホエノールピルビン酸カルボキシラーゼ(PEPCK)とGTPの作用でリン酸化されてホスホエノールピルビン酸に戻される。
- ホスホエノールピルビン酸は解糖と共通の酵素の働きで逆経路を進み、ATPを1分子使って1.3ビスホスホグリセリン酸になり、アルドール縮合でフルクトース1.6ビスリン酸まで戻る。
- フルクトース1.6ビスリン酸は解糖ではATPを消費する不可逆な反応であったが、逆経路では解糖と別の酵素(フルクトースビスホスファターゼ)と水を使って加水分解を行ってリン酸基を取り除き、フルクトース6-リン酸に戻される。ここの酵素が糖新生の律速酵素である。
- フルクトース6-リン酸は解糖と共通酵素で異化され、グルコース6-リン酸に戻される
- グルコース6-リン酸は肝臓と腎臓の小胞体膜にのみ存在するグルコース6-リン酸トランスロカーゼと呼ばれる輸送体により小胞体の内部へ移動し、解糖と別の酵素(グルコース6-ホスファターゼ)と水で加水分解されてグルコースに戻される。生じたグルコースは特異的なトランスポーターで細胞質へ運び出される。
- 2分子のピルビン酸を1分子のグルコースに糖新生する反応は、(2ピルビン酸+4ATP+2GTP(ATP)+2NADH+H2O → グルコース+4ADP+2GDP(ADP)+6Pi+2NAD+)となることから、ピルビン酸からグルコース1分子を作る糖新生では6ATP使用することがわかる。
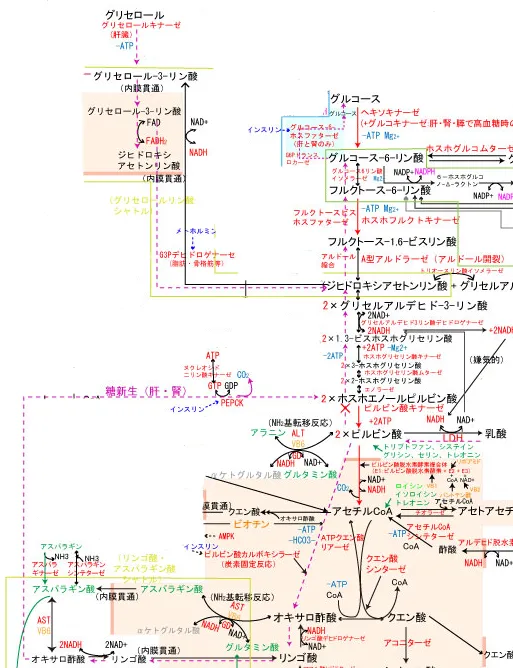
グリセロールからの糖新生
グリセロールからのルートは肝臓のグリセロキナーゼでグリセロール3-リン酸になった後にグリセロールリン酸シャトルの一部を形成し、ジヒドロキシアセトンリン酸へと変換される。
その後は共通の経路をたどりグルコースへと変換される。
グリコーゲンからの糖新生
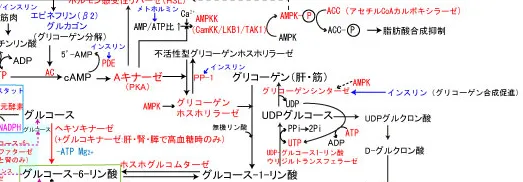
グリコーゲンの合成
過剰な糖質は、グルコース6-リン酸からホスホグルコムターゼの作用でグルコース1-リン酸に合成(可逆的な異性化反応)される。
グルコース1-リン酸はUTPによって高エネルギーのUDPグルコースへと活性化された後、グリコーゲンシンターゼの作用で既存のグリコーゲンの非還元末端に新たなグルコースを繋ぐ。
この時不要になったUDPが遊離するが、ヌクレオシド二リン酸キナーゼの作用でATPにリン酸化されてUTPに戻されて再利用される(G6P+ATP+グリコーゲン(n)+H2O → グリコーゲン(n+1)+ADP+2Pi)。反応式からもわかる通り、グリコーゲンの合成にはATPが消費される。
グリコーゲンの分解(糖新生)
肝臓のグリコーゲンはグリコーゲンホスホリラーゼにより加リン酸分解を受けると、グルコース1-リン酸を遊離する。グルコース1-リン酸はグルコース6-リン酸に戻された後、肝臓でグルコースまで戻される。
筋肉のグリコーゲンはグルコース6-リン酸からグルコースに糖新生できないので、そのまま解糖系を降下してATPの産生に回される。
乳酸からの糖新生
乳酸はピルビン酸まで戻された後に、TCA回路に進むか糖新生に回るかを選択することができる。
酸素があればTCA回路と酸化的リン酸化を通じてATPを獲得することも可能だが、アセチルCoAやATPが十分にあればATPを産生する必要性がないので、糖新生でグルコースを作る方に回る。
酸素が少ない高強度の運動で乳酸は蓄積していく。
この乳酸は骨格筋内でピルビン酸に戻した後TCA回路に入れることもできるが、血液を介して他の組織へ運ばれてTCA回路に入れることや、肝臓でピルビン酸からオキサロ酢酸を経てグルコースに戻されて、再度骨格筋に戻って解糖系を動かす(コリ回路=乳酸-グルコース回路=糖新生)こともできる。
コリ回路では、ピルビン酸から6ATPを消費してグルコースを作って骨格筋に送っても、解糖系では4ATPしか獲得できないので収支はマイナスとなり、無酸素運動を糖新生を使って持続することはもちろんできないし、酸素がある場合は乳酸を糖新生するよりも酸化的リン酸化に回した方がATP獲得効率がいい。
それではATP獲得効率が悪くても、なぜコリ回路が必要なのかというと、血糖のホメオスタシス(維持)です。
エネルギーとして乳酸を利用しているだけでは血糖が回復しないため、低強度の運動におけるコリ回路の役割は下がった血糖を維持する目的がある。
一方、高強度の運動ではグリコーゲンが枯渇しているような状態に陥っているため、血糖値の維持に加えて、グリコーゲンを再生するのにも役立っている。
血糖値を維持することは、ほとんどがグルコースの利用に頼っている脳(脳はケトン体も利用できる)や赤血球のような組織の機能を維持するためでもあります。
乳酸からの糖新生ではNAD+が必要となるが、メトホルミンはグリセロール3-リン酸デヒドロゲナーゼ2を阻害してグリセロールリン酸シャトルを阻害、結果、NAD+の供給が止められるため、糖新生は抑制される。
多量のアルコールも、エタノール→アセトアルデヒド→酢酸に変換されるときに、ADH(アルコール脱水素酵素)やALDH(アルデヒド脱水素酵素)とともに、NAD+が必要になるので、枯渇し、糖新生が抑制され低血糖と乳酸の増加を引き起こす。
糖原生アミノ酸からの糖新生、アミノ基転移反応
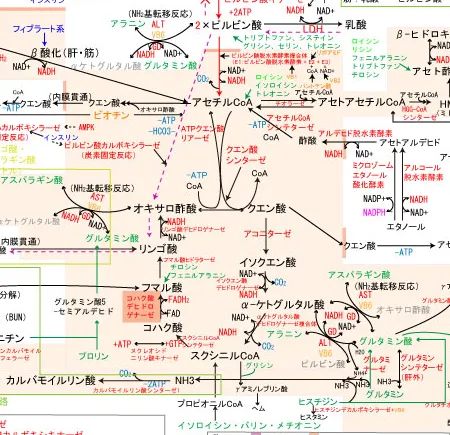
ロイシンとリシン以外は糖原生アミノ酸であり、糖新生の原料になります。
- トリプトファン、システイン、グリシン、セリン、トレオニンは代謝されるとピルビン酸に変換されるので、糖新生が可能
- アラニンはALT(アラニンアミノトランスフェラーゼ)とビタミンB6の元、アミノ基転移反応でαケトグルタル酸にアミノ基を渡してピルビン酸に変換されるので糖新生可能
- グルタミンとヒスチジン、プロリン、アルギニンはグルタミン酸になった後、GD(グルタミン酸デヒドロゲナーゼ)とNAD+の元、アミノ基転移反応でαケトグルタル酸へと変換されてTCA回路に組み込まれ、リンゴ酸から糖新生経路に入る
- イソロイシン、バリン、メチオニンはプロピオニルCoA→スクニシルCoAへと代謝されてTCA回路に組み込まれる
- チロシン、フェニルアラニンはフマル酸に代謝されてTCAサイクルに組み込まれる
- アスパラギン酸、アスパラギンはアミノ基転移反応でオキサロ酢酸へと変換されるので糖新生可能
なお、ケト原生アミノ酸もアセチルCoAに代謝されてTCA回路に入ることでリンゴ酸経由で糖新生可能に見えるが、そうはならない。
アセチルCoAが増えるとピルビン酸カルボキシラーゼが活性化されてピルビン酸からオキサロ酢酸へのルートが活性化するので、オキサロ酢酸が糖新生に優先的に回ってしまって、アセチルCoAと縮合できるオキサロ酢酸が不足し、アセチルCoAがTCA回路に入ることができなくなるためである。
アミノ基転移反応と酸化的脱アミノ反応
アミノ基転移反応とは、アミノ酸(アラニンやアスパラギン酸等)のアミノ基をαケト酸(αケトグルタル酸)に渡す反応である。
アミノトランスフェラーゼ(アラニンはアラニンアミノトランスフェラーゼ:ALT、アスパラギン酸はアスパラギン酸アミノトランスフェラーゼ:AST)とビタミンV6のリン酸化体(ピリドキサールリン酸)によって触媒される。
アミノ基を渡した側のアラニンやアスパラギン酸は対応するαケト酸(ピルビン酸やオキサロ酢酸)になり、渡された側のαケト酸(αケトグルタル酸)はグルタミン酸になる。
糖新生の基質となるピルビン酸やオキサロ酢酸を生成することが目的なので主に糖新生可能な肝臓や腎臓で行われる反応である。
アミノ基転移反応で任意のアミノ酸がアミノ基を転移する相手は必ずαケトグルタル酸であり、必ずグルタミン酸が生成するするのでグルタミン酸が生体内で最も多いアミノ酸になっている。
そして、このグルタミン酸をαケトグルタル酸に戻す反応が酸化的脱アミノ反応である。
酸化的脱アミノ反応はグルタミン酸をグルタミン酸デヒドロゲナーゼ(GD)とNAD+(NADP+でもOK)でαケトグルタル酸へと変換する過程で、アミノ基の窒素原子をアンモニアとして放出し、尿素回路(オルニチン回路)に回す。
酸化的脱アミノ反応は、アミノ基転移反応と並行して起こる反応ではあるが、必ずしも糖新生に並行して起こる反応ではないことが重要である。
酸化的脱アミノ反応は肝臓以外の組織でも起こっており、グルタミン酸に集められたアミノ基をアンモニアとして放出しているが、尿素回路は肝臓に存在する(肝臓に尿素回路に必要な2つの酵素が存在している)ため、末梢組織で発生したアンモニアをグルタミンシンテターゼのもとにグルタミン酸に結合してグルタミンという形にして血中を移動し、肝臓まで運んで、グルタミナーゼでグルタミン酸に戻す過程で末梢組織から取り込んだアンモニアを放出して肝臓の尿素回路に回す。
アミノ基転移反応は糖新生の基質であるピルビン酸とオキサロ酢酸をアミノ酸から作り出すことが重要であるため、並行してαケトグルタル酸が減少してTCA回路が回らなくなっても問題はない。
糖原生関連薬
糖新生に関連する医薬品は以下
参考サイト:薬学部学生向け講義資料ダウンロード
- 糖質の構造と種類
- 脂質の種類
- タンパク質の種類
-
ビタミン
├ 脂溶性ビタミン
| ├ ビタミンA
| ├ ビタミンD
| ├ ビタミンE
| ├ ビタミンF
| └ ビタミンK
|
└ 水溶性ビタミン
├ ビタミンB1
├ ビタミンB2
├ ビタミンB6
├ ビタミンB12
├ ビタミンC
├ ビタミンM
├ ビタミンH
├ ビタミンP
├ ビタミンQ
├ ニコチン酸
└ パントテン酸
コメントor補足情報orご指摘あればをお願いします。
- << 前のページ
- 次のページ >>
